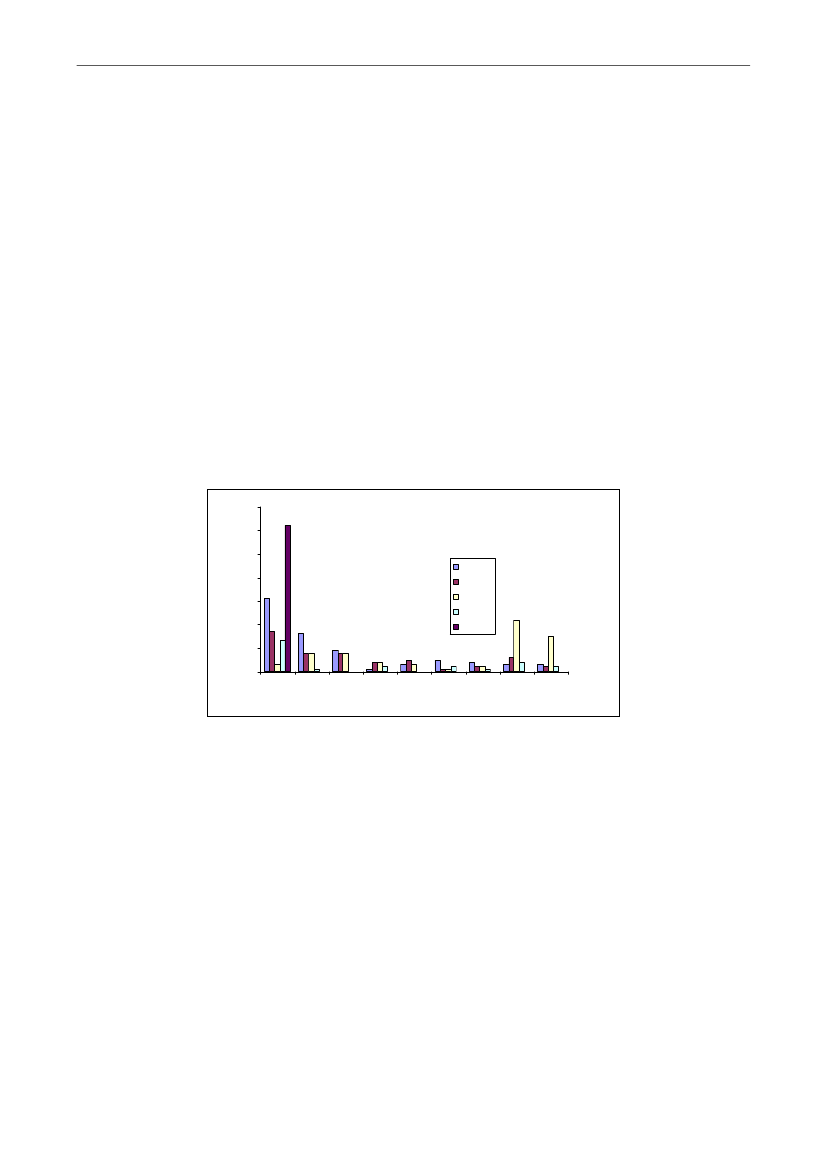Udvalget for Fødevarer, Landbrug og Fiskeri 2010-11 (1. samling)
FLF Alm.del Bilag 128
Offentligt
DET JORDBRUGSVIDEN-SKABELIGE FAKULTET
INSTITUT FOR HUSDYRBIOLOGIOG SUNDHED
PATTEGRISDØDELIGHED I DKMULIGHEDER FOR REDUKTION AF PATTEGRISEDØDELIGHEDEN I DANMARKPEER-REVIEWED INTERN RAPPORT
Lene Juul Pedersen,Peer BergErik JørgensenMarianne Kjær BondeMette S. HerskinKristian Møllegaard Knage-RasmussenAnne Grete KongstedCharlotte LauridsenNiels OksbjergHanne Damgaard PoulsenDaniel Alberto SorensenGuosheng SuMartin Tang SørensenPeter Kappel TheilKaren ThodbergKarin Hjelholt JensenInstitut for Husdyrbiologi og SundhedDet Jordbrugsvidenskabelige FakultetAarhus UniversitetBlichers Allé 20Postboks 508830 Tjele
Interne rapporter indeholder hovedsagelig forskningsresultaterog forsøgsopgørelser som primært henvender sig til DJF medarbej-dere og samarbejdspartnere. Rapporterne kan ligeledes fungeresom bilag til temamøder. Rapporterne kan også beskrive interneforhold og retningslinier for DJF .
Publikationer fra Det JordbrugsvidenskabeligeFakultet kan downloades på www.agrsci.au.dk
Tryk: www.digisource.dk
PATTEGRISDØDELIGHED I DK
Forord .......................................................................................................................3Sammendrag ............................................................................................................ 5Summary ................................................................................................................... 6KAPITEL 11.11.21.31.41.5FAKTA OM PATTEGRISEDØDELIGHED I DANMARK .......................................................... 8
UDVIKLING SIDEN1992I PATTEGRISEDØDELIGHED................................................................................ 8UDVIKLING GENNEM DE SENESTE20ÅR I AVL RELATERET TIL REPRODUKTION.......................................... 9UDVIKLING GENNEM DE SENESTE20ÅR I OPSTALDNING I FARESTALDEN OG DRÆGTIGHEDSSTALDEN...... 12PATTEGRISEDØDELIGHED I UDLANDET................................................................................................. 12SAMMENFATNING............................................................................................................................... 15
KAPITEL 2HVAD DØR GRISENE AF OG HVILKE EGENSKABER VED GRISENE DISPONERER FORDØDELIGHED?............................................................................................................................................... 162.12.22.32.42.52.6TOTAL DØDELIGHED........................................................................................................................... 16DØDFØDTE GRISE.............................................................................................................................. 17IHJELLÆGNING................................................................................................................................... 18SULT OG KULDE................................................................................................................................. 19SYGDOM............................................................................................................................................ 19VELFÆRDSMÆSSIGE KONSEKVENSER FOR DE DØENDE GRISE............................................................... 20
2.6.12.6.22.6.32.6.42.6.52.7
Dødfødte..................................................................................................................................21Kulde........................................................................................................................................21Sult...........................................................................................................................................22Sygdom/skader........................................................................................................................22Kvælning..................................................................................................................................22
SAMMENFATNING............................................................................................................................... 22STRATEGIER TIL REDUKTION AF TIDLIG PATTEGRISEDØDELIGHED .......................... 24
KAPITEL 33.1
FØDSEL AF FÆRDIGUDVIKLEDE GRISE MED HØJ VITALITET OG AF PASSENDE STØRRELSE........................ 24
3.1.13.1.23.1.33.23.3
Avlforfødselsvægtogoverlevelse..........................................................................................24Fødselafstørreogmerelevedygtigegrisegennemfodringafsoensidstidrægtighed........27Fødselafstørregriseogmerelevedygtigegrisegennemforbedretmanagement................29
SIKRING AF AT GRISEN ER I STAND TIL HURTIGT AT KOMME TIL PATTERNE OG INDTAGE RÅMÆLK.............. 32SIKRING AF GRISENS MULIGHED FOR AT INDTAGE RÅMÆLK/MÆLK......................................................... 32
3.3.13.3.23.3.33.4
Betydningafstimiljøforpattegriseadgangtilogstimulationafpatterne.............................33Brugafammesøer...................................................................................................................34Avldersikrer,atkuldstørrelseertilpassetantalpatter..........................................................36Energiindholdogmængdeafråmælk.....................................................................................38
SØER DER PRODUCERER TILSTRÆKKELIGT RÅMÆLK/MÆLK AF HØJ KVALITET......................................... 37
3.4.1
1
PATTEGRISDØDELIGHED I DK
3.4.23.4.33.4.43.5
Immunologiskkvalitetafråmælk............................................................................................39Forebyggelseaffarefeber .......................................................................................................40.Håndteringaffarefeber...........................................................................................................41Udvælgelseafsøermedgodemoderegenskaber...................................................................41Stimuleringafsoensmoderegenskabervedtildelingafpladsoghalm..................................42Forebyggelseafsygdomhossoen...........................................................................................44Reduktionafsygdomhosgrisene ...........................................................................................45.
PÅPASSELIGE SØER OG KVIKKE GRISE................................................................................................. 41
3.5.13.5.23.5.33.5.43.6
SAMMENFATNING............................................................................................................................... 45POTENTIELLE TILTAG, DER KAN REDUCERE PATTEGRISEDØDELIGHEDEN ............. 47KONKLUSION ......................................................................................................................... 50
KAPITEL 4KAPITEL 5
Referencer .............................................................................................................. 51Ordliste .................................................................................................................... 73Bilag ........................................................................................................................ 76
2
PATTEGRISDØDELIGHED I DK
ForordPå baggrund af den aktuelle debat i Danmark om dødeligheden af pattegrise i forhold tilsøernesøgedekuldstørrelseharFødevarestyrelsenanmodetDetJordbrugsvidenskabelige Fakultet, Aarhus Universitet, om en peer reviewed redegørelseforfattet på dansk vedrørende pattegrisedødelighed.De centrale elementer i den danske debat har været, om det danske avlsarbejde for fødselaf større kuld af grise har ført til en øget pattegrisedødelighed, om avl med sådannekonsekvenser er etisk forsvarlig, i hvilket omfang problemstillingen også er etdyrevelfærdsmæssigt problem, og om ændringen i det danske avlsmål fra kuldstørrelse til”levende grise dag 5” i 2004 har nedbragt pattegrisedødeligheden.På baggrund af den danske debat har Fødevarestyrelsen derfor ønsket en redegørelse forden foreliggende viden om årsager til de seneste års udvikling i pattegrisedødelighed iDanmark og i andre EU-lande samt en vurdering af hvilke metoder, der kan reducereproblemet. Desuden har man ønsket en redegørelse for velfærden hos ammesøer, dadisse er en konsekvens af, at soen føder flere grise, end den har patter til. Endelig harman ønsket en vurdering af velfærden hos grise, der dør.Disse emner behandles i nærværende rapport. Idet størstedelen af pattegrisene dørindenfor de første uger efter fødsel, har vi fokuseret på problemstillinger knyttet til dettetidsrum.Rapporten er udarbejdet i et tværdisciplinært samarbejde mellem forskere i genetik,etologi, ernæring, immunologi, kødkvalitet, management og beslutningsstøtte.SeniorforskerPeer Berg,SeniorforskerGuosheng Su,ForskningsprofessorDaniel AlbertoSorensenog ForsøgslederErik Jørgensen,alle fra Institut for Genetik & Bioteknologi, harbidraget med input og diskussion af de avlsmæssige aspekter og beregningsmæssigeovervejelser.SeniorforskerKaren Thodberg,Institut for Husdyrbiologi og -sundhed, har bidraget medmulighederne for at avle for forbedrede moderegenskaber hos søerne.SeniorforskerMette S. Herskin,Institut for Husdyrbiologi og -sundhed, har bidraget medafsnittet om lidelse hos døende grise.SeniorforskerMarianne Kjær Bondeog Ph.d.-studerendeKristian Møllegaard Knage-Rasmussen,Institut for Husdyrbiologi og -sundhed, samt ForskerAnne Grete Kongsted,Institut for Jordbrugsproduktion og Miljø, har bidraget indenfor overvejelser omkringmanagement.SeniorforskerCharlotte Lauridsen,ForskningslederHanne Damgaard Poulsen,SeniorforskerMartin Tang Sørensen,og SeniorforskerPeter Kappel Theil,alle fra Institutfor Husdyrbiologi og -sundhed, samt ForskningsprofessorNiels Oksbjerg,Institut forFødevarekvalitet, har leveret input til de ernæringsmæssige problemstillinger.3
PATTEGRISDØDELIGHED I DK
SeniorforskerLene Juul Pedersenog SeniorforskerKarin Hjelholt Jensenhar bidragetfagligt inden for deres ekspertiseområder og har forestået revidering af rapporten.Desuden har Marianne Kjær Bonde og Tina Albertsen læst korrektur og Mette GravesMadsen har forestået opsætning af rapporten. Endelig har Michael Groes Christiansen,Flemming Thorup og Poul Bækbo, Videncenter for Svineproduktion, bidraget med atfremskaffe materiale, der ikke umiddelbart er offentligt tilgængeligt.Rapporten er gransket af professor Lotta Rydhmer, Institut för husdjursgenetik, Sverigeslantbruksuniversitet, Uppsala, og professor Bo Algers, Institutionen för husdjurens miljöoch hälsa, Sveriges Lantbruksuniversitet, Skara.Lene Juul Pedersen og Karin Hjelholt Jensen, oktober 2010.
4
PATTEGRISDØDELIGHED I DK
SammendragRapporten gennemgår fakta om udvikling i Danmark i pattegrisedødelighed, udvikling iavlsarbejdet knyttet til reproduktion samt udvikling i staldsystemer til søer i de seneste årog sammenligner den nuværende pattegrisedødelighed i Danmark med tilsvarende tal fraudlandet. De seneste års avl mod større kuldstørrelse er formodentlig en stor del afårsagen til den stigende andel af både dødfødte grise og grise, der dør efter fødsel, iperioden fra 1992 til 2004. Andelen af dødfødte grise er dog formodentlig overvurderet,mens andelen af grise, der dør efter fødsel er undervurderet. Den stigende hyppighed afskift fra løsdrift i drægtighedsstalden til fiksering i farestalden, der er sket isvineproduktionen i samme tidsrum, kan i mindre omfang have bidraget til en øgetpattegrisedødelighed. Efter inddragelse af antal levende grise ved dag 5 som avlsmål istedet for kuldstørrelse er stigningen i andelen af døde grise stagneret, men der er ikke setdet forventede fald. Sammenligning af data vedrørende pattegrisedødelighed fra udlandetog Danmark tyder på, at danske søer har den markant største kuldstørrelse og totaledødelighed. Den højere dødelighed ser ud til primært at skyldes et højere antal, der erdødfødte eller dør indenfor de første timer efter fødslen. Den danske pattegrisedødelighedefter det første halve levedøgn er lavere end i nogle og større end i andre af de lande medintensiv svineproduktion, som vi har data fra.Den foreliggende viden om de biologiske forhold, der er afgørende for pattegrisenesoverlevelse, viser, at store kuldstørrelser fører til, at flere grise fødes små, svage ellerunderudviklede, og disse grise har øget risiko for at dø enten under eller efter fødslen.Dødeligheden efter fødsel er forårsaget af ihjellægning, sult/kulde og sygdom, og dissedødsårsager optræder med næsten samme gennemsnitlige hyppighed. Den relativebetydning af de forskellige typer af pattegrisedødelighed varierer dog mellem besætninger.Sammenfattende er store kuld, lav fødselsvægt, underafkøling og manglenderåmælksoptagelse i de første levetimer betydende risikofaktorer for grisenes overlevelse.De grise som dør efter fødslen udsættes sandsynligvis for en væsentlig lidelse varende frafå minutter op til et halvt døgn. Den høje andel af døde grise er derfor både et etisk ogvelfærdsmæssigt problem. Den store kuldstørrelse har desuden nødvendiggjort brug afammesøer for at forhindre at overskydende grise dør ved deres biologiske mor som følgeaf manglende patter til alle grise. Brug af ammesøer udgør dog et velfærdsmæssigtproblem for ammesoen, fordi det betyder, at den fikseres i en længere periode.Mulige strategier til reduktion af pattegrisedødeligheden gennemgås og vurderes irapporten. Inkludering af pattegriseoverlevelse i det danske avlsindeks er en oplagtmulighed for at reducere pattegrisedødelighed, og de nødvendige oplysninger ogberegninger synes allerede tilgængelige eller kan tilvejebringes med begrænset indsats.Dette vil dog ikke umiddelbart føre til en reduktion af pattegrisedødelighed iproduktionsbesætningerne, da det tager nogle år, før avlsarbejdet slår igennem. Derimodhar en øget faringsovervågning potentiale for nu og her at reducere både andelen afdødfødte grise og andelen af grise, der dør efter faring, ligesom forbedringer i grisenes5
PATTEGRISDØDELIGHED I DK
nærmiljø i det omfang, der er udviklet praktisk anvendelige metoder til det, vil kunnereducere dødeligheden. Intensiveret rådgivning omkring sosundhed har ligeledes etpotentiale for, indenfor en kortere årrække, at reducere andelen af døde grise. På sigt vilflere andre tiltag kunne vise sig at have en positiv indflydelse, men disse forudsætteryderligere forskning, udvikling eller afprøvning.
SummaryThis report examines the development in Denmark in piglet mortality and in the breedingwork related to reproduction and housing systems for sows in recent years. Further itcompares the current piglet mortality in Denmark with similar figures from abroad. Thebreeding towards larger litter size carried out since 1992 and until 2004 is probably a largepart of the reason for the growing rates of both stillborn piglets and piglets that die afterbirth. There are reasons to believe that the proportion of stillborn piglets given in theannual reports from the pig industry is overvalued, while the proportion of piglets that dieafter birth is underestimated. During the same period there has been an increase in thenumber of farms where the sows switch from pens in the gestation barn to crating in thefarrowing barn. This switch causes stress in the sow and may to a lesser extent havecontributed to an increase in piglet mortality. After including number of live piglets at day 5in the Danish breeding index instead of litter size the increase in the proportion of deadpiglets stagnated, but adecreasein the piglet mortality has not been observed althoughthis was expected. Comparison of data on piglet mortality from abroad and Denmarkindicates that Danish sows have by far the largest litter size and the largest total mortality.The larger mortality seems mainly to be due to a larger rate of stillborn piglets or pigletsthat die within the first hours after birth. After this period the Danish piglet mortality is lowerthan in some and higher than in the other countries with intensive pig production fromwhich we have data.The present understanding of the biological factors that are crucial for piglet survivalshows that large litter sizes result in more piglets being born small, weak orunderdeveloped. These piglets have an increased risk of dying during or after birth.Mortality after birth is caused by crushing, starvation and cold and disease. These causesof death occur with almost the same average frequency. However, the relative importanceof the different types of piglet mortality varies between herds. In summary, large litters, lowbirth weight, hypothermia and lack of colostrum uptake in the first hours of life aresignificant risk factors for piglet survival.Those piglets that die after birth are probably exposed to a significant suffering lasting froma few minutes up to half a day. The high proportion of dead piglets is therefore both anethical and welfare problem. The large litter size has also necessitated the use of nursingsows. This has become necessary in order to prevent excess piglets to die by theirbiological mother due to lack of teats to all piglets. The use of nursing sows, however,constitutes a welfare problem for the nursing sow, because it means that the period ofcrating will be prolonged.6
PATTEGRISDØDELIGHED I DK
Possible strategies for reduction of piglet mortality are examined and estimated in thisreport. Inclusion of piglet survival in the Danish breeding index is one opportunity to reducepiglet mortality. The necessary information and calculations seem already available or canbe obtained with little effort. However, including the mortality rate in the breeding index willnot lead to an immediate reduction of piglet mortality in the production as it takes someyears before breeding takes place. Therefore, there will still be a need for improvedmanagement to reduce the current mortality. One way that has proven efficient to reduceboth the number of still born piglets and mortality after birth is to increase birth surveillanceand birth assistance to the new born piglets and sow. In addition improvement of themicroclimate at the birth site of the piglets may also have the potential to reduce mortality.However, there may be a need to develop practical solutions before these can beimplemented. Intensified advice about improvement of sow health may be another way toreduce production factors that are related to mortality. Eventually, several otherapproaches could prove to have a positive influence, but these require further research,development or testing.
7
PATTEGRISDØDELIGHED I DK
Kapitel 1Fakta ompattegrisedødelighed i Danmark1.1
Udvikling siden 1992 i pattegrisedødelighed
Den seneste årsopgørelse (2009) af pattegrisedødelighed viser, at der i gennemsnit i 2009blev født 16,1 grise pr. kuld, hvoraf 1,9 grise var dødfødte, svarende til 11,8 %, og 2 grisedøde efter fødsel og inden fravænning svarende til 12,4 % (1). Udviklingen iårsopgørelserne siden 1992 ses i Figur 1. Som det fremgår, er der sket en stigning fra1992 til 2009 både i antallet af fødte grise, i andelen af dødfødte grise og i andelen afdøde grise totalt set. Andel af grise, der dør efter fødsel er ligeledes steget, men ikke isamme omfang som de dødfødte grise.Opgørelserne er lavet af Videnscenter for Svineproduktion på basis af indberetning afproduktionsresultater til PC-baserede programmer til produktionsstyring, AgroSoft ogDLBR SvineiIT, også kaldet P-kontrol/E-kontrol-systemer. Indberetningerne sker ikke fraalle danske besætninger. For eksempel er der er en lille overvægt af indrapportering frastore besætninger, og opgørelserne kan ikke betragtes som fuldt ud repræsentative.Indtil 2008 omfattede resultaterne kun rådgivningscenterets eget system (nuværendenavn: DLBR SvineiIT). I perioden fra 2002 til 2008 var antallet af besætninger, som indgik iopgørelsen stærkt faldende med det laveste antal besætninger i 2007, hvor der kun indgik171 besætninger med et gennemsnitligt antal årssøer på 500. Perioden fra 1992 til 1996inkluderer ca. 15 % af alle besætninger, varierende fra 2237 besætninger medgennemsnitligt 154 årssøer i 1992 til 1825 besætninger med gennemsnitligt 168 årssøer i1995. I 2009 blev datagrundlaget udvidet med Agrosofts programløsning, og der indgik666 besætninger med gennemsnitlig 579 årssøer pr. besætning, svarende til ca. 35 % afde danske søer og ca. 20 % af alle besætninger med søer.I sådanne opgørelser er der en vis usikkerhed i hvor mange grise, der er dødfødte, oghvor mange grise, der dør efter faring. Produktionskontrolsystemer anvendes først ogfremmest i den daglige produktionstilrettelæggelse i besætningerne. Oftest vil de grise, derer døde, første gang søerne tilses efter faringens afslutning, blive registreret somdødfødte, selvom nogle af dem først er døde umiddelbart efter fødslen. Denvidenskabelige definition af dødfødte er, at de er døde under selve fødslen, men detkræver en obduktion af grisene for sikkert at konstatere, om de har trukket vejret efterfødslen. Det er derfor sandsynligt, at andelen af dødfødte grise er overvurderet i forhold tilden videnskabelige definition, og at andelen af grise, der først dør efter fødsel ertilsvarende undervurderet i de danske opgørelser (Figur 1). Under danskeproduktionsforhold sker der tillige en omfattende omfordeling af grise mellem søerumiddelbart efter faringen. Denne omfordeling bevirker yderligere fejlmuligheder iregistreringssystemet. Der kan være store forskelle mellem besætningerne i, hvordan
8
PATTEGRISDØDELIGHED I DK
registreringen af de enkelte produktionsegenskaber foregår og dermed i kvaliteten af deindsamlede data.
Udvikling i procent døde af total og antal fødte grise2523211917151311975Procent/antal grise
Dødfødte (%)Døde efter faring (%)Døde, ialt (%)Antal fødte, total
19
År
Figur 1. Udvikling siden 1992 i procent døde i alt, procent dødfødte grise og procent døde før fravænningblandt grise registreret som levendefødte, og antallet af fødte grise i kuldet. Procenttallene er alle udtrykt iforhold til det totale antal fødte grise i kuldet. Kurven er lavet ud fra tal opgivet i årsberetninger fraVidencenter for svineproduktion. (1).
1.2
Udvikling gennem de seneste 20 år i avl relateret tilreproduktion
Hovedparten af de grise, der produceres i Danmark, har baggrund i det danskeavlsprogram DanAvl, enten i form af at sopoltene indkøbes fra avls- elleropformeringsbesætninger, og/eller at der købes sæd fra KS-stationer, hvor der tappessæd fra avlsorner. Avlsbesætninger driver selektionsarbejdet på renracede dyr, mensopformeringsbesætningerne, producerer de krydsningsdyr, der fortrinsvis anvendes iproduktionsbesætningerne. Der kan købes grise fra andre avlsselskaber i Danmark. Disseavlsselskaber har deres avlsarbejde i andre lande, og er kun repræsenteret i Danmarkeller nabolandene med opformeringsbesætninger. I det følgende fokuseres på at beskrivedet danske avlssystem, dels fordi størstedelen af den danske svineproduktion er baseretpå avlsmateriale fra det danske avlssystem, og dels fordi DanAvl har en åben beskrivelseaf deres avlssystem i modsætning til de internationale avlsselskaber. DanAvladministreres af Videncenter for Svineproduktion (VSP). Dermed er DanAvl de danskesvineproducenters eget avlssystem.
9
20
08 06/2009
94
98
00
19
93
96
19
20
20
20
02
92/
19
20
19
04
PATTEGRISDØDELIGHED I DK
Avlen er baseret på, at der beregnes avlsværdier for de enkelte egenskaber ud fraregistreringer på dyret selv og dets slægtninge. Disse avlsværdier vægtes sammen medden økonomiske vægt for de enkelte egenskaber, i en samlet avlsværdi(selektionsindeks). Forudsætningerne for beregning af denne samlede avlsværdi erkendskab til egenskabernes økonomiske værdi, arvbarhed samt genetiske korrelationmellem egenskaber. Den økonomiske vægt vælges, så den repræsenterer den marginaleøkonomiske effekt af at ændre det genetiske niveau for egenskaben én enhed. I Tabel 1er de egenskaber, der indgår i avlsarbejdet i 2009, præsenteret sammen med deresøkonomiske vægt og avlsfremgangen for de enkelte egenskaber i 2009. Heraf fremgår, atlevende grise ved dag 5 (LG5) er én af flere egenskaber, der i dag indgår i avlsmålet. LG5indgår kun i avlsmålet for hundyrracerne Landrace og Yorkshire, da det erkrydsningsafkom fra disse to racer, der anvendes som produktionssøer i sobesætninger.Duroc racen anvendes derimod udelukkende som handyrrace til krydsningsafkom, derender som slagtesvin. Derfor lægges der primært vægt på avl for tilvækst ogfoderudnyttelse i denne race. Ændringer i avlsværdien for egenskaber, der ikke indgår iindekset vægtes implicit med en økonomisk værdi på 0. Det er her vigtigt at forstå, at deter forholdet mellem de økonomiske vægte, der har betydning. Hvis den økonomiske vægtfor en egenskab øges, vil det svække den relative vægt for andre egenskaber og dervedsvække den forventede avlsfremgang på disse, fordi dens relative økonomiske værdivurderes som lavere. En øget fremgang for kuldstørrelse vil for eksempel ske påbekostning af daglig tilvækst. Effekten af at inkludere en ny egenskab i indekset kan altsåikke vurderes isoleret for egenskaben, men skal ses i sammenhæng med konsekvensenpå avlsfremgangen for de øvrige egenskaber. En (negativ) økonomisk vægt på dødelighedvil svække fremgang i kuldstørrelse, men styrke fremgang i daglig tilvækst, på grund af degenetiske korrelationer. Hvis de økonomiske vægte værdisættes korrekt, sikrerselektionsindekset imidlertid, at avlsfremgangen giver størst mulig økonomisk fremgang. Idet danske avlsarbejde vil det sige størst fremgang per produceret kilo kød.
EgenskabTilvækst (30-100 kg)FoderudnyttelseKødprocentLG5 (levende grise dag 5)BenstyrkeSoens holdbarhedTilvækst (0-30 kg)
Økonomisk vægt. kr.0,12-1328,6013**12,5*42,5**0,12
Avlsfremgang10,6 g/dag0,030 FEs/kg0,16 %0,4 gris**0,03 point*-0,021 %**1,4 g/dag
10
PATTEGRISDØDELIGHED I DK
Slagtesvind* kun Duroc; ** kun Landrace og Yorkshire
-5
-0,014 kg/gris
Tabel 1. Økonomiske vægte (udtrykt som værdi per enhedsændring) og avlsfremgang per år i det danskeavlssystem i 2009. Kilde: Videncenter for Svineproduktion, 2010 (2).
Igennem de seneste 20 år er der sket væsentlige ændringer i avlsprogrammerne medhensyn til søers reproduktion, herunder pattegrisedødelighed.I 1980’erne var de fleste avlsprogrammer designet til at øge produktionsegenskaber somdaglig tilvækst, foderudnyttelse og kødprocent. I starten af halvfemserne begyndte man atinteressere sig for reproduktionsegenskaber. Selvom det var kendt, at frugtbarhed er ensammensat egenskab, blev kuldstørrelse den egenskab, man inddrog somselektionskriterium for at øge reproduktionen. Dette skyldtes dels, at kuldstørrelse er nemat registrere og dels, at en del studier pegede på, at kuldstørrelse var den egenskab, somhavde den største økonomiske betydning for soens reproduktionsværdi (f.eks. 3).Imidlertid var der i halvfemserne begrænset belæg for, at det var muligt at øgekuldstørrelse ved direkte selektion (4), bortset fra resultaterne fra projektet,”hyperprolificdesigns” (5-7). Nye beregningsmetoder og større computerkraft antydede dog, at det varmuligt.På denne baggrund blev kuldstørrelse i 1993 indført i avlsmålet i det danske avlsprogramfor racerne, Landrace og Yorkshire, ved at tilføje kuldstørrelse som en af egenskaberne iselektionsindekset. Kuldstørrelse blev målt som det totale antal fødte grise pr. kuld.Resultaterne af det ændrede avlsprogram blev evalueret i en undersøgelse sidst i1990'erne. Denne viste meget tydeligt, at kuldstørrelse kunne øges ved genetisk selektion(8). Imidlertid viste undersøgelsen også tydeligt, at en øget kuldstørrelse var ledsaget afen lavere fødselsvægt med større variation inden for kuld og en større andel af dødfødtegrise pr. kuld.Disse ugunstige forhold i form af øget dødelighed og lavere fødselsvægt blev baggrundenfor projektet ”SuperSo” i 2002, hvor målet var at undersøge alternative metoder til at øgekuldstørrelsen og samtidig kontrollere andelen af dødfødte grise pr. kuld. I lighed med dentidligere undersøgelse (8) fandtes igen at den genetiske korrelation mellem det totale antalfødte grise pr. kuld og grisenes overlevelsesevne er negativ, mens den genetiskekorrelation mellem kuldstørrelse ved dag 5 (LG5) og andel overlevende dag 5 er svagtpositiv (9). De estimerede korrelationer tydede på, at man ved at selektere for LG5 villeopnå et svagt fald i andelen af døde grise. Dette begrundede, at kuldstørrelse kunneerstattes af kuldstørrelse ved dag 5 uden at øge antallet af egenskaber iselektionsindekset og derved svække selektionsfremgangen for de øvrigeproduktionsegenskaber såsom tilvækst. Estimaterne af de genetiske parametre i de todanske soracer, Landrace og Yorkshire, gav anledning til en forventning om en øgetpattegriseoverlevelse på 2 % ved en stigning i kuldstørrelse med 1 gris, hvilket ville11
PATTEGRISDØDELIGHED I DK
resultere i et svagt fald i den absolutte dødelighed som følge af selektion for LG5 (9). Somresultat af projektet blev LG5 inddraget i avlsmålet i 2004 som mål for kuldstørrelse, istedet for den oprindelige egenskab, som var det totale antal fødte grise pr. kuld. LG5registreres og evalueres som en egenskab hos soen. Smågrisens genetiske kapacitet foroverlevelse antages ikke at påvirke antal levende grise ved dag 5.Som det fremgår af Figur 1, er der ikke sket den forventede reduktion i andelen af dødegrise i produktionsbesætningerne, men snarere en stagnering. Derimod er den forventedefremgang i kuldstørrelse bekræftet.
1.3
Udvikling gennem de seneste 20 år i opstaldning ifarestalden og drægtighedsstalden
Der foreligger ikke en egentlig årlig opgørelse over, hvordan danske søer opstaldes. Ifarestalden vurderer vi, at ca. 99 % af søerne fikseres i såkaldte kassestier fra indsættelseca. én uge før faring til fravænning af kuldet ca. fire uger efter faring. En opgørelse afstistørrelser i 84 danske besætninger i 2008 (Bilag 1) viste, at bredden af farestiernevarierede mellem 115 og 248 cm, at længden af farestierne varierede mellem 200 og 315cm, og at arealet af farestien varierede mellem 2,85 og 6,15 m2. Vi vurderer også, at derikke er sket væsentlige ændringer i kassestiens størrelse gennem de seneste 20 år. Dogkom der en anbefaling fra Videncenter For Svineproduktion (10) om at øge kassestiernesdimensioner til 4,9 m2da det var vist at søernes størrelse var øget væsentligt gennem deseneste 15-20 år. I drægtighedsstaldene er der gennem de seneste 20 år sket envæsentlig ændring, idet en stor del besætninger er overgået fra fiksering af drægtige søertil gruppeopstaldning grundet EU’s krav om gruppeopstaldning fra 4 uger efter fravænningog indtil 1 uge før faring. Der er ikke helt overenstemmelse mellem, hvad forskellige kilderangiver som status for hvor stor en andel af besætningerne, der er overgået til løsdrift.Andersen et al. (2010) angiver at 68 % af de drægtige søer er løsgående (11), mens enanden angiver, at kun 55-61 % af søerne i dag er løsgående (12). Ændringen iopstaldningsforholdene kan have bidraget til stigningen i andelen af dødfødte grise. Ensvensk undersøgelse har vist, at opstaldning i løsdrift under drægtigheden efterfulgt affiksering i farestalden fører til en øget andel af dødfødte grise, især hos gylte (13).Implementeringen af løsdrift i drægtigheden er dog ikke sket jævnt over årene, og kanderfor ikke alene forklare den gradvise stigning i antallet af dødfødte grise, som ses påFigur 1.
1.4
Pattegrisedødelighed i udlandet
De fleste avlsselskaber, der leverer svin til de lande, der har intensiv svineproduktion, harligesom Danmark inddraget avl for større kuld i avlsarbejdet. Kun få avlsselskaber angiver,12
PATTEGRISDØDELIGHED I DK
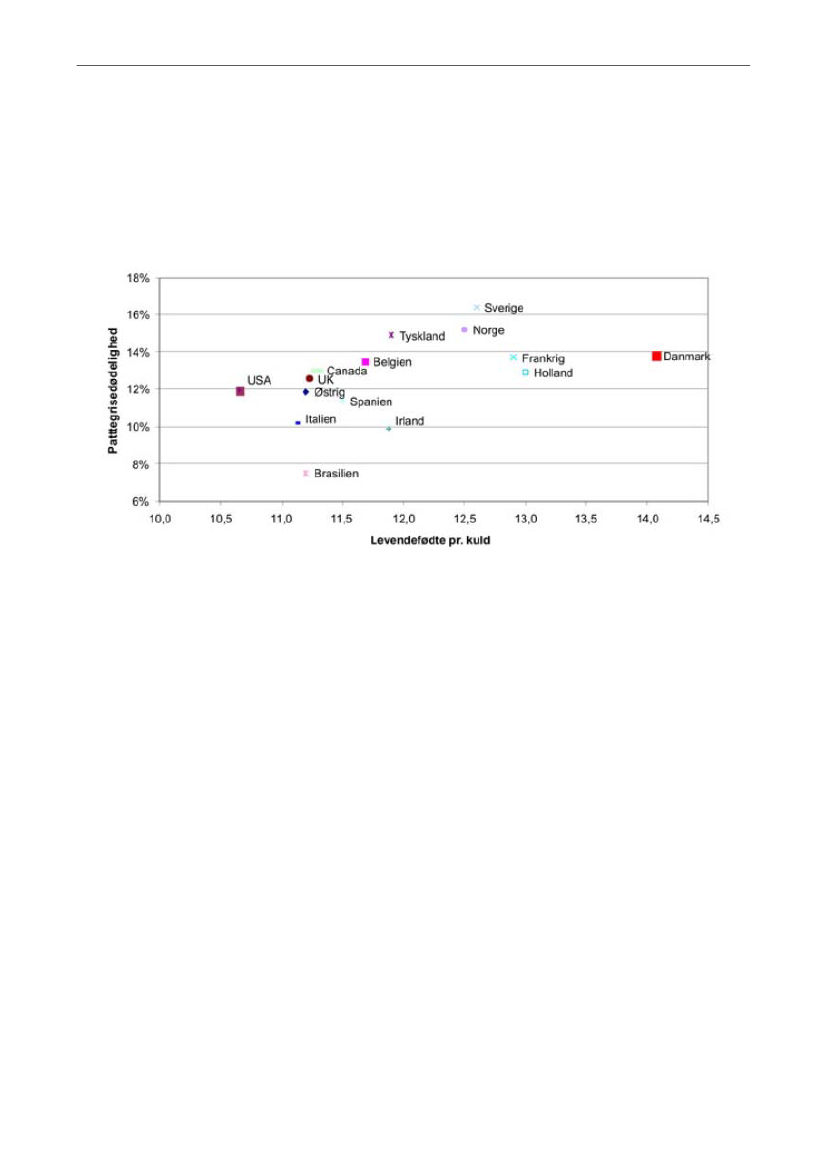
at aspekter relateret til overlevelse indgår i deres avlsmål (14). Avlsarbejdet i udlandetsynes imidlertid at have været mindre effektivt, set i forhold til kuldstørrelse, end iDanmark. De danske søer har en markant større kuldstørrelse end de andre lande, vi harkunnet finde oplysninger fra. På Figur 2 ses andelen af døde grise blandt de levendefødtei forhold til antallet af levendefødte grise.
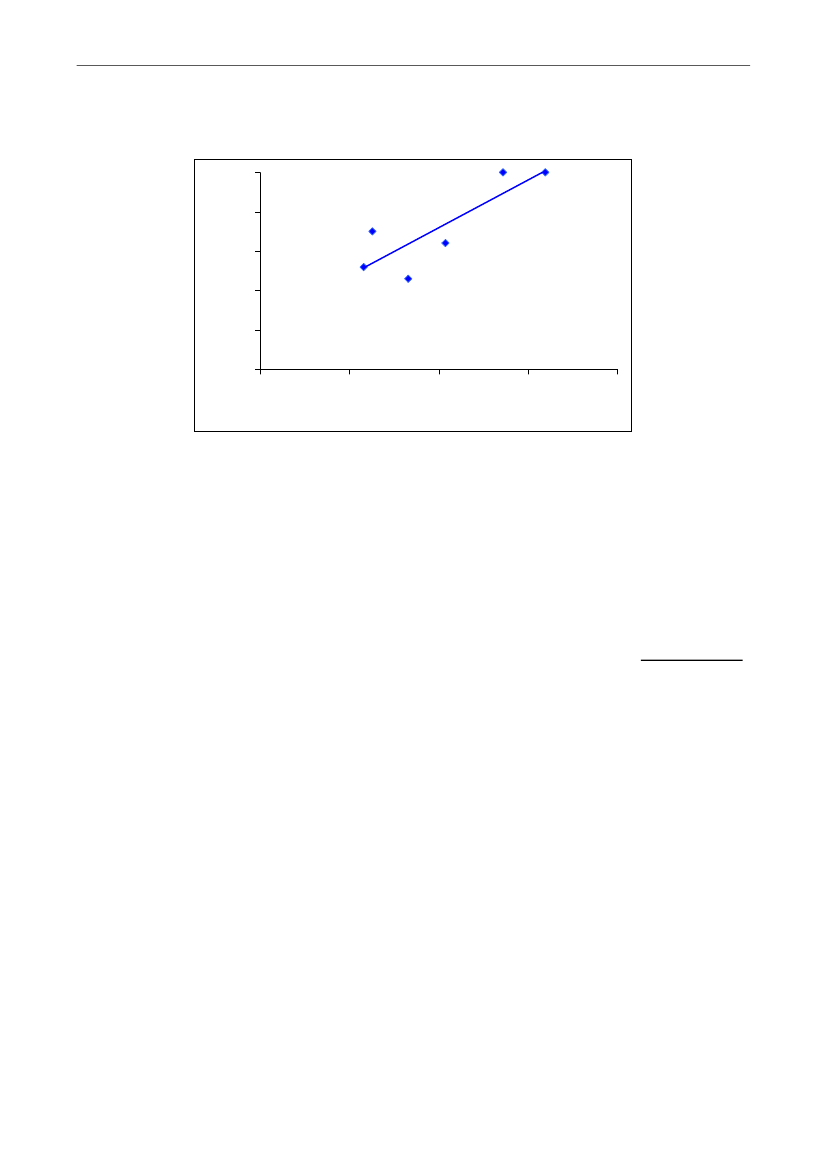
Figur 2. Levendefødte pr. kuld & pattegrisedødelighed. De viste data stammer fra indberetninger til InterPIG,der er en international sammenslutning mellem en række europæiske lande samt Brasilien, USA og Canadaog har til formål at benchmarke produktionsøkonomien i svineproduktionen mellem de deltagende lande.Kilde: Christiansen, 2010 (15)
Som det fremgår, får danske søer i gennemsnit over en levendefødt gris mere end søernei andre lande. Med hensyn til den registrerede dødelighed af de levendefødte pattegriseligger Danmark i den øvre ende, men overgås af lande, som Tyskland, Sverige og Norge.Som nævnt i afsnit 1.1. er tallene for antallet af levende grise sandsynligvis undervurderetpå grund af registreringsusikkerheden på, om døde grise har været dødfødte eller først erdøde kort tid efter fødslen. Hvor meget denne usikkerhed påvirker tallene kan væreforskelligt mellem lande. Andelen af døde grise blandt de levendefødte grise varierede i2008 med 0,6 gris mellem gode og dårlige besætninger i Danmark (1), hvilket viser, atdenne del af pattegrisedødeligheden i høj grad afhænger af managementfaktorer ogregistreringspraksis.Blandt de lande, hvor det har været muligt at fremskaffe data på antallet af totalt fødtegrise og den totale pattegrisedødelighed, er den totale pattegrisedødelighed mellem 0,9 og3,7 % højere i Danmark end i de øvrige lande (Figur 3).
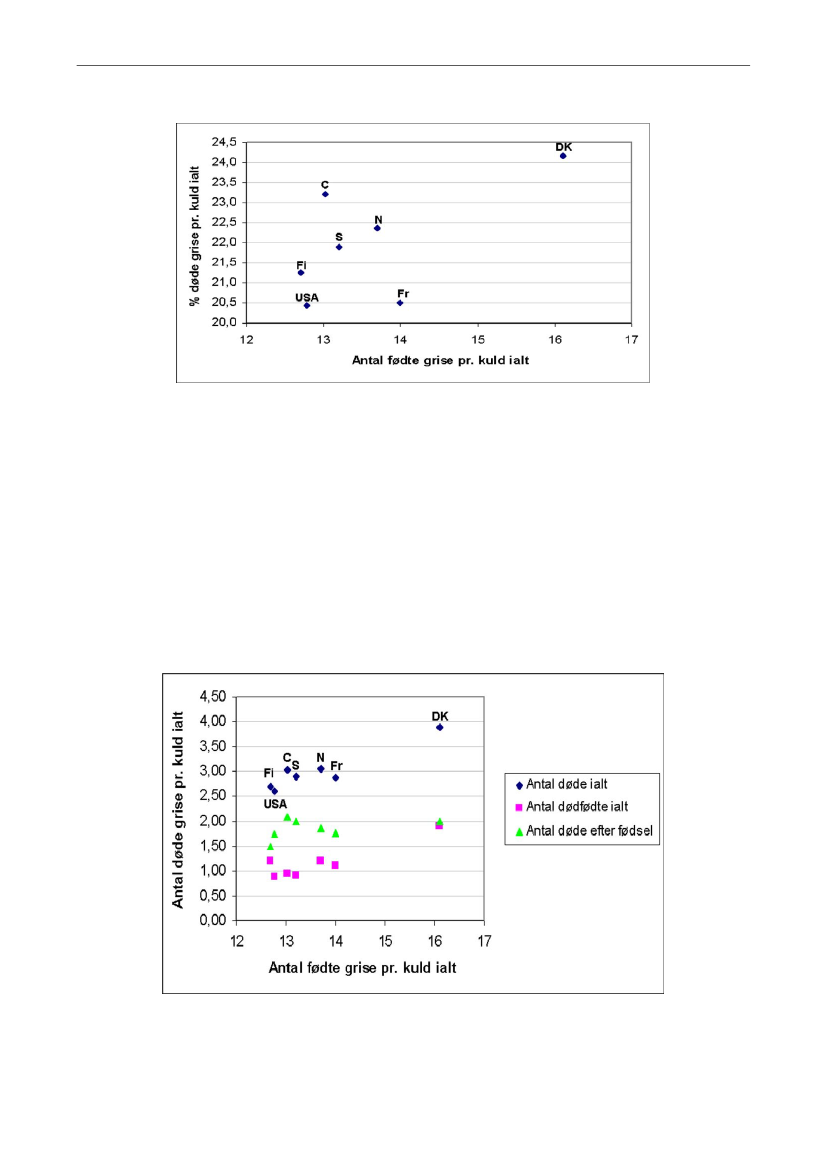
13
PATTEGRISDØDELIGHED I DK
Figur 3. Andelen af døde grise i alt i forhold til det totale antal fødte grise i Finland (Fi), Sverige (S) og Norge(N) i 2007 (16-18), i Frankrig (Fr) i 2008 (19), i Danmark (DK) i 2009 (1), og i USA og Canada (C) i 2009 (20).
Som det fremgår af Figur 4, er der på tværs af lande en tydelig sammenhæng mellem,hvor mange grise søerne får, og hvor mange grise der registreres som dødfødte, dvs. hvormange grise, der enten er dødfødte eller dør tidligt efter fødslen. Antallet af registrerededødfødte grise er 0,7 grise større i Danmark end i Norge og Finland, der er de lande, derellers har det største antal dødfødte grise. I overensstemmelse med at de registrerededødfødte grise i høj grad er bestemt af kuldstørrelsen er variationen mellem gode ogdårlige besætninger i antallet af dødfødte grise kun 0,1 gris i danske besætninger (1).Antallet af grise der dør efter fødslen viser i disse opgørelser ikke klare sammenhængemed kuldstørrelsen.
14
PATTEGRISDØDELIGHED I DK
Figur 4. Antallet af dødfødte grise, døde levendefødte grise og døde grise i alt i forhold til det totale antalfødte grise i Finland (Fi), Sverige (S) og Norge (N) i 2007 (16-18), i Frankrig (Fr) i 2008 (19), i Danmark (DK)i 2009 (1), og i USA og Canada (C) i 2009 (20).
Sammenligning af data fra udlandet med de danske data tyder samlet set på, at danskesøer har den markant største kuldstørrelse både ved faring og ved fravænning, men atomkostningen er, at flere grise er dødfødte eller dør kort tid efter fødslen. Der er ikkemulighed for at vurdere, hvor stor andel af de registrerede dødfødte grise, der først dørefter fødslen, og hvis velfærd efter al sandsynlighed har været kompromitteret jf. afsnit 2.6.1.5
Sammenfatning
De seneste års avl mod større kuldstørrelse synes at udgøre en stor del af årsagen til denstigende andel af både dødfødte grise, og grise, der dør efter fødsel i perioden fra 1992 til2004. Andelen af dødfødte grise er dog formodentlig overvurderet, mens andelen af grise,der dør efter fødsel, er undervurderet. Den stigende hyppighed af skift fra løsdrift idrægtighedsstalden til fiksering i farestalden, der er sket i svineproduktionen i sammetidsrum, kan i mindre omfang have bidraget til en øget andel af dødfødte grise. Efterinddragelse af antal levende grise ved dag 5 som avlsmål i stedet for kuldstørrelse, erstigningen i andelen af døde grise stagneret, men der er ikke set det forventede fald.Sammenligning af data vedrørende pattegrisedødelighed fra udlandet og Danmark tyderpå, at danske søer har den markant største kuldstørrelse, og at flere grise er dødfødteeller dør indenfor de første timer efter fødslen. Den danske pattegrisedødelighed efter detførste halve levedøgn er lavere end i nogle og større end i andre af de lande med intensivsvineproduktion, som vi har data fra.Produktionsbesætningerne står derfor med et avlsmateriale, som formodentlig levererverdens højeste kuldstørrelse, men desværre også har en høj dødelighed, som bådeudgør et økonomisk tab for landmanden og samtidig udgør et velfærdsmæssigt og etiskproblem.
15
PATTEGRISDØDELIGHED I DK
Kapitel 2
Hvad dør grisene af oghvilke egenskaber ved grisenedisponerer for dødelighed?
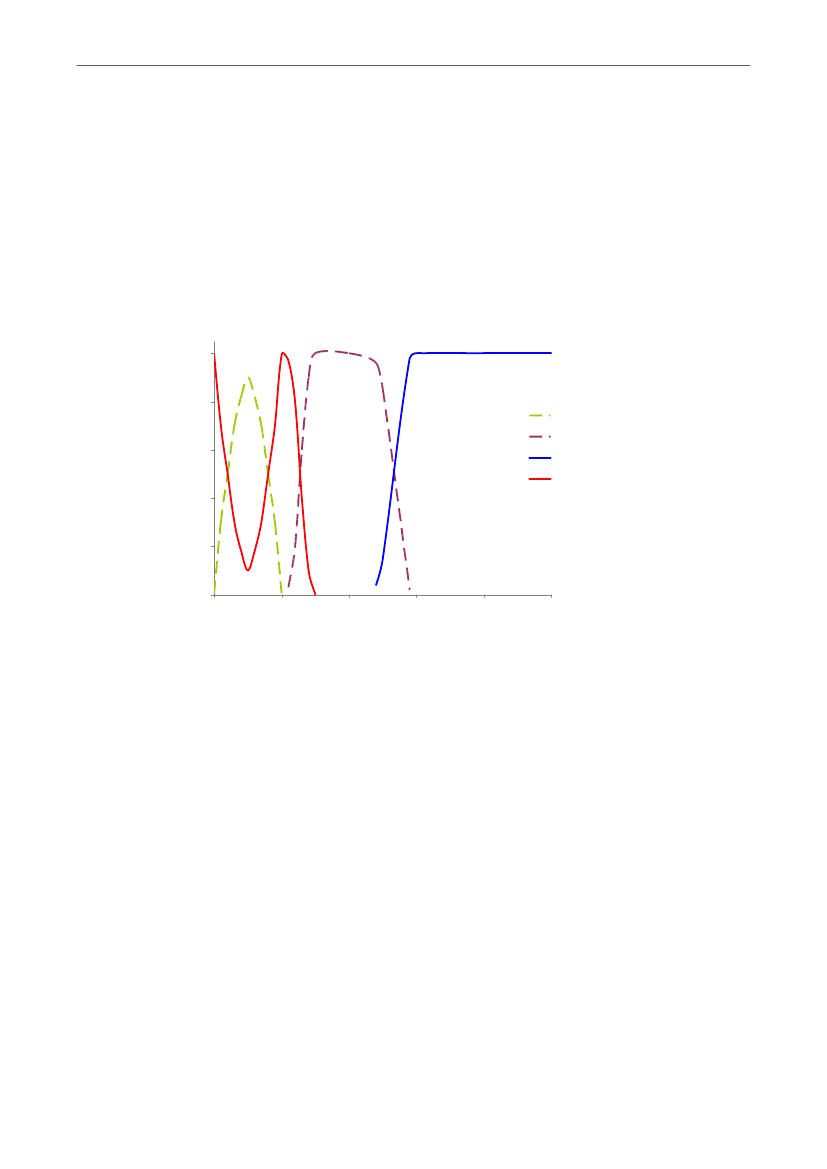
Hovedparten af pattegrisene dør i de første levedage, hvilket bl.a. fremgår af Figur 5, somer obduktionsdata fra en dansk undersøgelse, hvor døde grise fra 87 faringer er obduceretog sammenholdt med video (21). Den tidsmæssige udvikling er i overensstemmelse medandre danske og internationale undersøgelser (f.eks. 9, 22-24).Den totale pattegrisedødelighed kan tilskrives flere typer af pattegrisedød. Nogle grise dørunder fødslen, nogle lægges ihjel af soen eller dør af sult eller kulde, mens andre igen døraf sygdom. I det følgende beskrives de biologiske forhold, der er afgørende for deforskellige typer af pattegrisedød.
7060number of piglets50crushed4030201000102030405060708+15+day after birthstarveddiseaseotherstillborn
Figur 5. Udviklingen gennem diegivningsperioden i antal grise der dør (i alt 302) af forskellige årsager i endansk undersøgelse, hvor døde pattegrise fra 87 faringer (halvdelen fra kassestier og halvdelen fraløsdriftsstier) blev obduceret og sammenholdt med video for at bekræfte dødsårsagen (21).
2.1
Total dødelighed
Der er en klar og veldokumenteret sammenhæng mellem kuldstørrelse og andel af dødegrise i kuldet, idet flere fødte grise totalt medfører øget dødelighed både i traditionellekassestier (f.eks. 25-30), i løsdriftstier (31, 32) og under udeproduktion (33). Årsagen erblandt andet, at en forøgelse i kuldstørrelsen forårsages af en øget ovulationsrate ogdermed implantering af flere fostre i den tidlige drægtighed. Mange fostre i den tidligedrægtighed resulterer i dårligere vækstforhold for de enkelte fostre, og nogle fostre går til
16
PATTEGRISDØDELIGHED I DK
grunde undervejs pga. enten pladsmangel eller underernæring. Uanset hvor mange grisesoen ender med at føde, kan flere eller alle grise i et kuld være påvirket negativt af defostre, der gik til grunde, fordi de påvirkede udviklingen af placenta (34). Nedsat vækst ogudvikling af embryoner/fostre eller deres organer betegnes Intra Uterin Growth Retardation(IUGR). I grise kan IURG være forårsaget af soens under- eller overernæring,varmestress, sygdom eller toxin indtagelse (35). Underernæring af fostrene kan ogsåopstå som følge af en begrænset kapacitet i livmoderen, hvor blod- og næringsstoftilførseler utilstrækkelig til et ’overskud’ af fostre. En del af de underernærede fostre dør længe førfødsel, mens andre ikke er tilstrækkeligt udviklet på fødselstidspunktet og derfor dør ligefør eller under fødslen. Andre fødes små og underudviklede (30, 36) med en forøget risikofor at dø (30). Detaljerede undersøgelser af døde grises organer viser blandt andetforandringer i organvægt, hormonelle forstyrrelser og ændringer i binyrebarken hos degrise, der er døde omkring fødslen (30). En række undersøgelser viser også en klarnegativ sammenhæng mellem kuldstørrelse og fødselsvægt (21, 37-39). Et stort antalfostre i tidlig drægtighed resulterer generelt i en lavere gennemsnitlig fødselsvægt og enstørre variation i fødselsvægt indenfor kuldet (34). Forsøg med direkte selektion forintrauterin kapacitet antyder, at øget intrauterin kapacitet øger overlevelsen af fostre (40)primært mellem d 25 og 45 i drægtigheden og kan erkendes indtil 10 dage før faring. Derer dog ingen oplysninger om overlevelse efter fødsel da alle gylte blev aflivet før faring.Som det fremgår af nedenstående er fødselsvægt også af afgørende betydning forpattegrisenes overlevelse, når der ses på de enkelte dødsårsager.
2.2
Dødfødte grise
Selve fødslen er én af de største udfordringer for den endnu ufødte gris, og andelen afdødfødte grise udgør ca. halvdelen af tabet af pattegrise i soproduktionen. Hovedparten(mere end 70 %) af de grise, som er dødfødte, synes at være døde som følge afhændelser under selve faringen (41). Ifølge dataopgørelser fra de besætninger, somindrapporterer til P-kontrollen i Danmark, er 11 % af alle grise dødfødte (kapitel 1). Iavlsbesætninger med renracede søer rapporterer Su et al. (2007), at andelen af dødfødtegrise er 15,6 % (9), mens udenlandske undersøgelser angiver, at 4-8 % af grisene erdødfødte (42), dog med et lavere totalt antal fødte grise, hvilket gør sammenligningvanskelig. Opgørelser fra besætninger kan som tidligere nævnt imidlertid være behæftetmed fejl, idet ikke vitale grise, der dør kort efter faringen, fejlagtigt kan bedømmes somdødfødte af landmanden. At der findes en sådan skævvridning i den subjektivebedømmelse af dødsårsager bekræftes af flere undersøgelser, hvor landmandens egenvurdering er sammenlignet med obduktionsfund (41, 43-45). Fra en nylig afsluttet danskundersøgelse omfattende 87 kuld, hvoraf halvdelen kom fra løse søer og halvdelen frasøer i kassestier, blev der på baggrund af obduktion fundet 5,2 % dødfødte grise (21) (ikkeforskellig mellem løsdrift og kassestier). Selvom der i undersøgelsen blev ydetfaringshjælp, hvis fødselsintervallet mellem grisene oversteg 3 timer, støtterundersøgelsen, at den høje andel af dødfødte grise, som findes via P-rapportens
17
PATTEGRISDØDELIGHED I DK
opgørelse, omfatter både dødfødte grise og grise, som er døde i de første levetimeromkring soen. Dødfødte grise har ofte været udsat for iltmangel under fødslen som følgeaf lang tids ophold i fødselskanalen (21, 31, 46-50). Forlænget ophold i fødselskanalenudgør ikke kun en risiko for dødfødte grise, men øger også de levendefødte grises risikofor at dø af andre årsager (f.eks. sygdom, dårlig trivsel o. lign.) (31).En del undersøgelser peger på, at tunge grise har en lavere risiko for at være dødfødteend lettere grise (21, 50, 51, 52), men der er også fundet genetiske sammenhængemellem højere fødselsvægt og øget frekvens af dødfødte (52). Pattegrise med lavfødselsvægt har ofte været underernærede gennem fosterudviklingen pga. en dårligerefungerende placenta under drægtigheden og fødes derfor som nævnt både undervægtigeog ofte underudviklede. Disse grise kan være i større risiko for, at navlestrengen brydesinden fødsel, hvorved de udsættes for en højere risiko for iltmangel under faringen. Detteunderstøttes af, at laktat indholdet i grisenes blod, der er udtryk for graden af iltmangel, erhøjere hos levende nyfødte grise med lav fødselsvægt end hos grise med højerefødselsvægt (21). Desuden er det vist, at blodets evne til at transportere oxygen varringere hos grise med lav fødselsvægt end hos grise med høj fødselsvægt (53). Ligeledeshar undersøgelser vist at grise med lav fødselsvægt også ofte har underviklede organer ogfødes med mindre fysiologisk modning og forstørrede binyrer (30). Der synes således atvære tale om at grise, som fødes små og fra store kuld ikke alene er mere udsat for at døpga. deres størrelse men i lige så høj grad fordi de fødes umodne.Mange undersøgelser har desuden vist, at øget kuldstørrelse disponerer for øget risiko fordødfødte grise (25, 26, 31, 48, 54, 55), hvilket sandsynligvis forårsages af den tidligereomtalte effekt af kuldstørrelse på fostrenes udvikling og vækst.
2.3
Ihjellægning
En del døde grise er klemt af soen og er enten døde af dette eller har været døde, førsoen lagde sig på dem. Flere undersøgelser tyder på, at ihjellægning tæller istørrelsesordenen 5 % af totalt fødte grise.Opgørelser fra renracede søer fra danske avlsbesætninger viser, at 3,3 % af grisene erklassificeret af landmanden som værende lagt ihjel af soen (9). En anden danskundersøgelse i en enkelt besætning fandt en lidt højere andel af ihjellagte grise (5,8 % aftotal fødte grise). I de 5,8 % var inkluderet både små undervægtige grise uden mælk imaven såvel som grise, hvor ihjellægning var den eneste forklarende årsag til død (21). I(9)fandtesudoverde3,3 % ihjellagte desuden, at ca. 2,9 % af det totale antal fødte grise blev kategoriseret som”tidlig død” (9), hvilket er grise, som er døde inden landmandens første observation afkuldet. En del af disse grise er sandsynligvis også klemte. Udenlandske undersøgelserbekræfter, at ca. 5 % af alle fødte grise dør som følge af ihjellægning (24), men igen er detvanskeligt at sammenligne til udenlandske undersøgelser, fordi kuldstørrelsen er
18
PATTEGRISDØDELIGHED I DK
væsentlig højere i DanmarK og derfor har en betragtelig indflydelse på andelen af dødegrise.De to vigtigste risikofaktorer for ihjellægning synes at være lav fødselsvægt og lavkropstemperatur 1-2 timer efter fødsel (21, 36, 50, 56, 57). Pattegrise, som mister varmetidligt efter fødslen, vil være mindre vitale og derved have sværere ved at undgå soensbevægelser. Grise med lav fødselsvægt vil have mere behov for næring og vil derfor væretilbøjelige til både at være mere aktive i længere tid omkring diegivningerne ved soensyver og at klumpe omkring yveret i hvileperioder. Herved udsætter de sig samtidig for enstørre risiko for at blive klemt af soens bevægelser (58, 59). Samtidig er små grise mereudsat for at miste varme, idet forholdet mellem deres overflade areal og kropsvægt erhøjere end store grises. Kulde nedsætter grisenes evne til at optage antistoffer overtarmen (60), og kan derfor føre til øget sygdomsmodtagelighed.2.4
Sult og kulde
En anden væsentlig kilde til pattegrisedødelighed er sult og kulde. I en danskundersøgelse udgjorde grise, som døde uden mælk i maven og uden tegn på skader frasoen eller sygdom 3,8 % af det totale antal fødte grise (21). Ca. 1/3 af de grise, som dødeuden mælk i maven, havde aldrig indtaget råmælk. Som for ihjellægning var det en lidthøjere andel end fundet i (9), hvilket igen kan skyldes, at kategorierne ”tidlig død” ogdødfødte kan have omfattet grise, der ikke havde fået mælk eller var døde af kulde (9).Det er vanskeligt at adskille sult og kulde i to dødsårsager. Ofte fører manglenderåmælksoptagelse til underafkøling, eller tidlig underafkøling svækker grisen i en sådangrad, at den ikke er i stand til at finde frem til patten og optage råmælk. De typiskerisikofaktorer hos grisen er igen lav fødselsvægt og lav kropstemperatur 1-2 timer efterfødsel (21, 50, 56). Ydermere udgør iltmangel under fødsel, f.eks. som følge af langefødselsintervaller (61), en risikofaktor (21, 62). Grise, som vurderet ud fra niveauet af laktati navlestrengsblodet har været udsat for iltmangel under fødslen, har sværere ved at findepatten, så de kan få råmælk (21, 61). Sen mælkeoptagelse kombineret med lav vitalitet ogevt. underafkøling, der tærer på grisenes egne energireserver i form af glykogendepoter,vil gøre det vanskeligt for den nyfødte gris at optage tilstrækkelig råmælk til at gøre denvital nok til at tilkæmpe sig en patte i det første levedøgn, hvor pattegrise etablerer derespatteorden (afsnit 3). Disse grise vil ikke nødvendigvis dø umiddelbart efter fødsel, menkan gradvist blive svagere, og have mere og mere vanskeligt ved at forsvare deres patteoverfor kuldsøskende og vil få mindre og mindre mælk. Af Figur 5 fremgår det, at flere afde grise, som dør uden mælk i maven, først dør 7 dage eller længere tid efter fødsel.2.5
Sygdom
Det er vanskeligt at angive et niveau for hvor mange pattegrise, der dør af sygdom, fordidet er meget afhængigt af den enkelte besætnings sundhedsstatus og det aktuellesmittepres. Opgørelse i 2010 fra den udvidede sundhedskontrol (USK) i danskebesætninger, hvor alle døde grise fra 10 tilfældigt udvalgte besætninger blev obduceret i19
PATTEGRISDØDELIGHED I DK
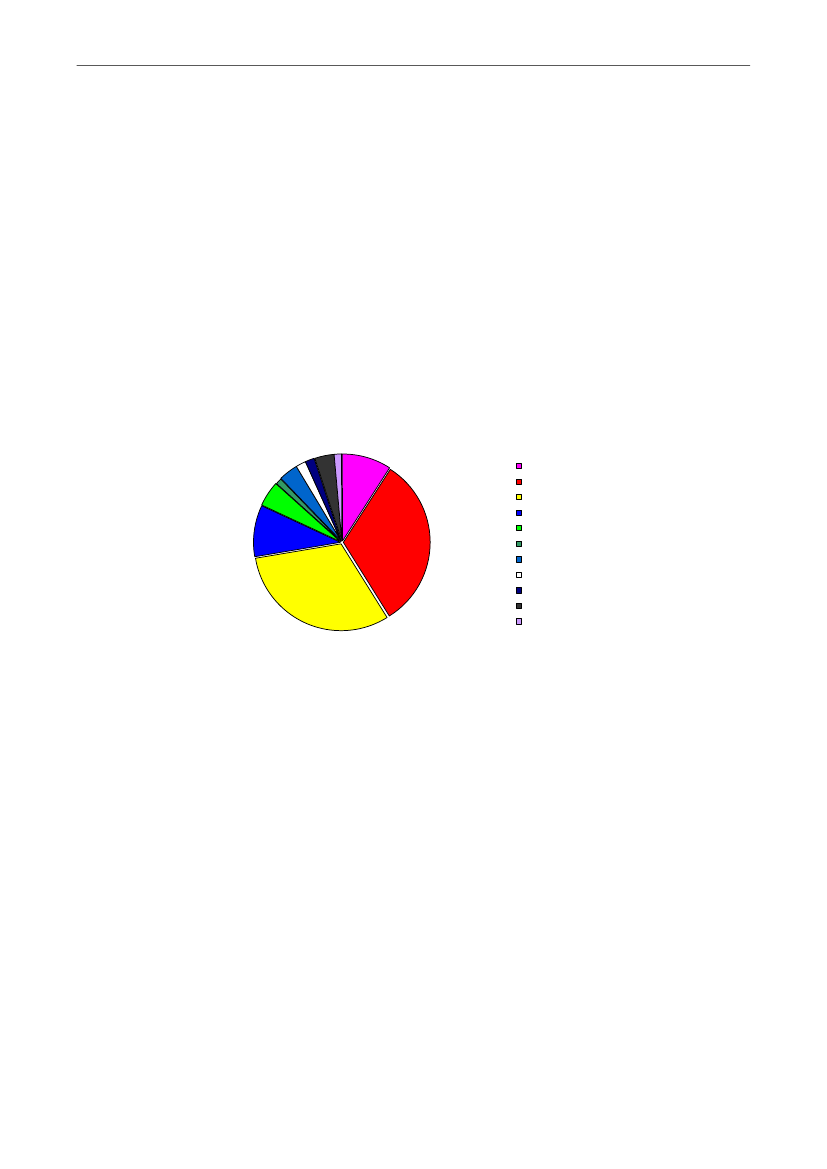
en periode, viser, at 22 % af de døde grise døde som følge af sygdom (primærtgeneraliseret infektion evt. med efterfølgende ledbetændelse, og diarré - se Figur 6). Dettesvarer til, at der i gennemsnit dør i størrelsesordenen 5 % af grisene af sygdom, ogniveauet synes uændret over de seneste 18 år (22). I en svensk undersøgelse i 12besætninger fra 2004 var diarré årsagen til 34 % af dødsfaldene indenfor de første 8 dageefter faring (63). Andelen af grise som døde af andet end ihjellægning var 6,3 % i en storSchweizisk besætningsundersøgelse (64).Som for de andre dødsårsager udgør fødselsvægt en væsentlig risikofaktor for, atpattegrise dør af sygdom (21, 65). Pedersen et al. (2010) fandt desuden, at lavkropstemperatur 2 timer efter fødsel også var disponerende for, om grisene døde afsygdom, hvilket i den pågældende undersøgelse primært var forårsaget af diarré (21 -Figur 5).Diagnoser på 718 pattegrise fra 10 USK-gris i 20102% 4%2%4%1%5%1%9%Undervægtig/ svagfødtKlemt / traumeSultSepticæmi/ arthritis10%31%EnteritisForstoppelseKastrationsskadeIntet specifikt fundOtitis mediaAndetPleuropneumoni31%
Figur 6. Data fra 10 tilfældigt udvalgte danske besætninger, hvor i alt 718 døde pattegrise er obduceret (22).Figuren viser fordelingen af diagnoser, som forårsagede døden hos de obducerede grise. Septicæmi/arthritiser blodforgiftning/ledbetændelse, enteritis er diarré, otitis media er mellemørebetændelse ogpleuropneumoni er lungebetændelse.
2.6
Velfærdsmæssige konsekvenser for de døende grise
Generelt er det en betingelse for tilstedeværelse af lidelse og reduceret dyrevelfærd, at etindivid er i live og bevidst nok til at erfare sensoriske input samt fortolke dem somubehagelige (66).Når dyr aflives hensigtsmæssigt (dvs. uden lidelse i form af smerter, frygt eller stressforbundet med proceduren (67), er der som sådan ikke velfærdsmæssige konsekvenserforbundet med døden. På trods af en betydelig indsats for at forebygge og forhindre tidligpattegrisedødelighed, har de velfærdsmæssige konsekvenser kun fået begrænsetopmærksomhed. Den eksisterende viden på området er samlet i et review fra 2004 (68),
20
PATTEGRISDØDELIGHED I DK
hvori de velfærdsmæssige konsekvenser ved med tidlig dødelighed hos de almindeligstehusdyrarter gennemgås.Man ved, at babyer – både efter normal graviditet og ved for tidlig fødsel – er i stand til atopfatte smerter umiddelbart efter fødslen. Viden om smerteopfattelse efter fødslen hos dyrer væsentligt mere begrænset (67), og hovedparten af den eksisterende viden stammerfra studier af rotteunger. Hos nyfødte pattegrise stammer næsten al viden omkringsmerteopfattelse fra studier af managementindgreb såsom halekupering eller kastration(se review af Herskin & Jensen, 2008). Her har en række studier vist at grise føler smerteved vævsskade selv ret tidligt efter fødslen (69), hvorfor vævsskade forud for en pattegris’død også vil være smertefuldt. Den præcise tidslige adskillelse mellem fosteretsmanglende smerteopfattelse og den nyfødtes evne til at opfatte smerter hænger sammenmed den fødsels-associerede ændring fra lav arteriel iltspænding i fosterstadiet til normalspænding efter fødslen. Den præcise timing i skiftet i smerteopfattelse fra foster til nyfødtgris kendes ikke, men da grise fødes højt udviklede sammenlignet med f.eks. rotter ellermennesker (de kan f.eks. gå omkring, se og koordinere adfærd med kuldsøskende fåtimer efter fødslen) er der intet grundlag for at forvente, at deres smerteopfattelse erringere end nyfødte babyers. Nedenfor gennemgås velfærdsmæssige konsekvenser vedde typiske dødsårsager hos pattegrise.
2.6.1 DødfødteDer findes ikke egentlige studier af pattegrises bevidsthedsniveau i fosterstadiet.Vurderingen må derfor foretages på baggrund af studier af fostre og nyfødte hosmennesker og andre pattedyr. Som nævnt tyder sådanne undersøgelser på, at fostrebefinder sig i en bevidsthedsmæssig dvaletilstand indtil de selv er i stand til at trækkevejret. Selvom de kan reagerer refleksmæssigt på stimuli, der efter fødslen ersmertevoldende, så viser de ikke tegn på, at de har en bevidst oplevelse af smerte, idetreaktionen ikke involverer centralnervesystemet, men kun er en perifer reaktion på stimuli(70). Der er derfor p.t. ingen dokumentation for, at fostre er bevidste om smerte elleranden stress, hvorfor dødfødte grise formodentlig ikke selv oplever reduceret velfærd. Detsamme gør sig gældende for dyr, der reelt er i live ved fødslen men dør umiddelbartderefter. Der er p.t. ikke data tilgængelige til at etablere en tidslig adskillelse fortidspunktet, hvor velfærdsproblemerne indfinder sig.
2.6.2 KuldeDer findes ikke studier, som har kortlagt den velfærdsmæssige betydning af afkøling forpattegrise. Fra mennesker og andre arter, ved man imidlertid at afkøling – med mindre detsker meget pludseligt, som ved fald i vand – ikke er en synderlig ubehagelig ellerstressende oplevelse. Mellor & Stafford (2004) beskriver, hvorledes et temperaturfald på3-7 grader relativt hurtigt efter et pattedyrs fødsel formodentlig vil føre til en betydeligreduktion i hjerne funktion (68). Hos pattegrise har man fundet, at et fald i kropstemperaturtil 34-35 grader fører til et fald i hjernens stofskifte og blodtilførsel på op til 40-50 % (71).Samlet tyder disse resultater på at underafkøling, som varer over ca. 30 minutter (grisekan leve i timevis efter en sådan nedkøling), ikke er forbundet med ubehag eller stress af21
PATTEGRISDØDELIGHED I DK
betydning, idet grisen gradvist glider over i en komalignende tilstand. I perioden førbevidsthedsreduktionen vil stressorer, såsom mas/klem fra so og kuldsøskende eller detikke at kunne følge sine søskende, dog kunne påvirke dyrets velfærd negativt.
2.6.3 SultNyfødte grise, som sultes, vil opleve sult i timerne før dødens indtræden, hvilket kanpåvirke deres velfærd negativt. Ofte vil der for grises vedkommende være tale om eninteraktion mellem sult og afkøling, som betyder, at sidstnævnte vil føre til omtaltebevidsthedsreduktion og deraf mindre påvirkning af grisens velfærd.
2.6.4 Sygdom/skaderCa. halvdelen af grise, der klemmes af soen, dør ikke umiddelbart, men kan leve videre itimer eller døgn efter, at skaden er sket. Disse dyr vil typisk have kvæstelser på indreorganer, knoglebrud og/eller sår. Typisk bliver disse grise aflivet, så snart landmandenopdager dem, og de lever således i op til 12-15 timer med skaden. I denne periode vilkvæstede dyr opleve smerter, hvis omfang afhænger af graden og typen af vævsskade.Sygdom vil pr. definition reducere dyrevelfærd, og syge dyr viser adfærdsmæssige tegn påubehag (72). Hvor påvirket det enkelte dyrs velfærd er, afhænger af typen og graden afsygdom.
2.6.5 KvælningHvis grisen kvæles i forbindelse med iltmangel under fødslen gælder – som for de reeltdødfødte – at de formodentlig ikke har nået et bevidsthedsniveau, der muliggør lidelse.Derfor er der sandsynligvis ikke det store velfærdsproblem forbundet med kvælning ifødselsøjeblikket.Hvis de derimod kvæles i forbindelse med ihjellægning efter selve fødslen, vil de være vedfuldt bevidsthed og være i stand til at opleve både frygt og stress i forbindelse medkvælningen. Man ved fra mennesker og andre dyr, at oplevelser af åndedrætsbesværudløser kraftige stress- og frygtreaktioner. Der findes kun meget begrænset viden omgrises stress- og frygtreaktioner i de første timer/dage af livet, men blandt andet på grundaf dyrenes høje grad af udvikling ved fødslen er der ikke belæg for at forvente, at de ikkekan føle stress eller frygt på dette tidspunkt. Blandt pattegrise, som dør ved ihjellægning,anslås det, at mindst halvdelen dør som følge af kvælning i løbet af 15-20 minutter. Ensådan langvarig kvælning under ihjellægning vil derfor have negative velfærdsmæssigekonsekvenser for dyrene.2.7
Sammenfatning
Grise, der er født små, svage eller underudviklet har øget risiko for at dø enten under ellerefter fødslen. Andelen af disse grise stiger med stigende kuldstørrelse. Blandt delevendefødte grise synes dødeligheden forårsaget af ihjellægning, sult/kulde og sygdom atvære af næsten samme størrelsesorden på nationalt plan. Den relative betydning af de22
PATTEGRISDØDELIGHED I DK
forskellige typer af pattegrisedødelighed vil variere mellem besætninger, ikke mindst medhensyn til sygdomsrelaterede dødsårsager. Sammenfattende er store kuld, lavfødselsvægt, underafkøling og manglende råmælksoptagelse i de første levetimerbetydende risikofaktorer for grisenes overlevelse.Hovedparten af de grise, som dør efter fødslen, udsættes sandsynligvis for en væsentliglidelse i form af enten smerte, sult, frygt eller stress varende fra få minutter op til et halvtdøgn.
23
PATTEGRISDØDELIGHED I DK
Kapitel 3
Strategier til reduktion aftidlig pattegrisedødelighed
Den stigning, der er set i kuldstørrelse og andel døde grise siden 1992, synes som nævntat være stagneret med indførelse af det nye avlsmål LG5, men det ser ikke ud til at haveresulteret i den forventede reduktion i andel døde grise. Både af etiske, velfærdsmæssigeog produktionsøkonomiske årsager er der derfor behov for at finde løsninger, der kanreducere pattegrisedødeligheden.Som det fremgår af ovenstående sker meget af dødeligheden i grisens første leveuge ogofte som følge af begivenheder, der enten er grundlagt i drægtighedsstadiet og/eller underfaringen og i grisens første levetimer. I det følgende vil vi derfor med udgangspunkt i deneksisterende viden gennemgå, hvilke muligheder der er for at fremme grisenesoverlevelse. Vi vil tage udgangspunkt i avl, staldindretning, fodring, sygdomsforebyggelseog management, der kan påvirke forhold af afgørende betydning for grisens overlevelse,dvs. fremme 1) at grisen fødes færdigudviklet med høj vitalitet og af en passendestørrelse, 2) at grisen er i stand til hurtigt at komme til patterne 3) at grisen er i stand til atindtage råmælk/mælk 4) at råmælken findes i tilstrækkelig mængde og kvalitet og 4) atsoen er påpasselig overfor grisene og grisene er kvikke, så de kan undgå uheldigebevægelser hos soen.
3.1
Fødsel af færdigudviklede grise med høj vitalitet og afpassende størrelse
3.1.1 Avl for fødselsvægt og overlevelseEt af de væsentligste forhold, der gør, at grisen er i stand til at klare sig igennem de førstekritiske levetimer uden varige men, er, at den fødes færdigudviklet og af en passendestørrelse. Fødselsvægt udgør således klart en grundlæggende faktor for pattegrisensoverlevelse (kapitel 2). Avlen for stigende kuldstørrelse har medvirket til, at der fødes enstor andel undervægtige og underudviklede grise, som det er beskrevet i afsnit 2.1.På nuværende tidspunkt er overlevelsesevne ikke direkte en del af selektionskriterierne idet danske avlsprogram. Inddragelsen af kuldstørrelse ved dag 5, der i 2004 erstattededet totale antal fødte grise per kuld i avlsindekset, var et skridt i den rigtige retning for atbegrænse stigningen i dødelighed, idet målet tager hensyn til, at der ikke bare fødesmange grise, men også at de overlever frem til dag 5. Kuldstørrelse ved dag 5 er dog ikkeet direkte udtryk for overlevelse og kan ikke forventes at give en markant øgetoverlevelsesprocent.
24
PATTEGRISDØDELIGHED I DK
Grisenes overlevelsesevne er påvirket af både moderens og grisens genotyper. Et effektivtselektionskriterium for at formindske dødelighed vil derfor være, at der selekteres bådedirekte for grisens egen evne til at overleve (direkte effekter) og for soens bidrag til at sikregrisene overlevelse (de maternelle genetiske effekter) (73). En mulighed for atimplementere dette er at selektere for maternelle genetiske effekter hos mor linjer og fordirekte effekter hos far linjer. Både direkte og maternelle genetiske effekter bør inddrages,dels fordi de begge bidrager, og dels fordi der i de fleste undersøgelser er fundet enugunstig genetisk sammenhæng mellem dem.Med henblik på at øge overlevelsesevnen for produktionsdyret (DxYL) undersøgerVidencenter for Svineproduktion i øjeblikket arveligheden for egenskaben ”overlevelse” tilslagtning (2). Her undersøges det, om der hos Duroc er arvelighed i overlevelsesevnenhos afkommet. Hvis det viser sig at være tilfældet, forventes egenskaben at kunneinddrages i avlsmålet.I hvor høj grad og hvor hurtigt inddragelse af selektion for overlevelse i det danskeavlsindeks vil kunne reducere pattegrisedødeligheden afhænger af, hvor stor økonomiskvægt egenskaben tillægges (jf. kapitel 1). En ny svensk undersøgelse simulerede etsimplificeret alvsprogram i én so-race med udgangspunkt i et konventionelt avlsindeks, derminder om det, der anvendes i Danmark (74). Avlsindekset inkluderer en økonomiskvægtning af de enkelte avlsegenskabers betydning og er således en økonomisk model.Undersøgelsen estimerede, at inkludering af både antal levende grise ved dag 5 ogoverlevelse fra dag 5 til fravænning i det nuværende avlsmål ville indebære en øgetkuldstørrelse (+0,11 gris) og en reduceret overlevelse fra dag 5 til fravænning (-0,1 %). Enfordobling af den økonomiske vægt på overlevelse ville føre til en konstant eller øgetoverlevelse og en reduceret stigning i kuldstørrelse, dog afhængig af den økonomiskevægt for de øvrige egenskaber i avlsmålet. Resultaterne er naturligvis afhængige af degenetiske parametre, der er antaget i denne undersøgelse, men de tyder på, at inkluderingaf overlevelse i avlsmålet vil have en gavnlig effekt på den genetiske udvikling ioverlevelse. Den genetiske ændring i overlevelse er dog følsom overfor de økonomiskevægtninger af egenskaber, der indgår i avlsmålet, og overfor de genetiske sammenhængemellem egenskaberne i avlsmålet.Endvidere viser undersøgelsen, at en øget vægtning af egenskaber, som overlevelse, vilresultere i en reduceret fremgang i produktionsegenskaber, som tilvækst og kødprocent.Hvis den økonomiske værdi af overlevelse tillægges tilstrækkelig betydning er der såledesikke noget til hinder for at opnå den ønskede udvikling. På nuværende tidspunkt er der idet danske avlsprogram ikke fastsat en økonomisk værdi af reduceret dødelighed, hvilketikke synes korrekt set ud fra både et produktionsøkonomisk, velfærdsmæssigt og etisksynspunkt. Det er f.eks. oplagt, at der er øgede omkostninger ved at fravænne 10 grise frakuld, hvor der blev født 20 grise, frem for at fravænne 10 grise fra kuld, hvor der blev født14 grise. Denne omkostning kan kun repræsenteres, hvis de ekstra døde grise har ennegativ økonomisk konsekvens i selektionsindekset. Med den nuværende vægtning afdødeligheden, som alene sker via LG5, vægtes antallet af døde grise ikke direkte iselektionen, men kun indirekte via den genetiske sammenhæng til LG5. Der må forventes25
PATTEGRISDØDELIGHED I DK
at være en vis omkostning for soen og de overlevende gris af antallet af døde grise, ogdette afspejles ikke i det nuværende alvsmål. Der kan derfor være god grund til at se på,hvordan de økonomiske vægte beregnes, herunder specielt tidshorisonten og hvilke typeraf omkostninger, der tages højde for. Her kan egenskaber som dødelighed, der giver etnegativt image, vægtes højere med en non-market værdi (75), der afspejler at grisekødkonkurrerer med andre kød typer om forbrugernes gunst.Grundet den klare fænotypiske sammenhæng mellem fødselsvægt og overlevelse (sekapitel 2) vil det desuden være nærliggende, at avl for større fødselsvægt vil kunnereducere andelen af døde grise. Mange undersøgelser viser, at der er en genetisksammenhæng mellem lavere fødselsvægt og reduceret kuldstørrelse (f.eks. 9, 33, 76, 77)og en ugunstig genetisk sammenhæng mellem fødselsvægt og overlevelse. Dette betyder,at selektion for store kuld må forventes at give lavere fødselsvægt, og lavere fødselsvægtkan forventes at give ringere overlevelse efter fødsel. Der er imidlertid også undersøgelseder viser at sammenhængen mellem fødselsvægt og overlevelse er forskellig afhængig afhvilken overlevelse man ser på. Der er f.eks. fundet negative sammenhænge mellemfødselsvægt og andel levendefødte (52). Den genetiske sammenhæng mellemfødselsvægt og overlevelse efter fødsel varierer fra undersøgelse til undersøgelse, menligger i størrelsesordenen 0,15 til 0,6 (33). Det betyder, at selektion for højere fødselsvægtsandsynligvis vil kunne reducere andelen af døde grise efter fødsel, men den relativt lavegenetiske sammenhæng mellem fødselsvægt og overlevelse indikerer, at det alt andet ligevil være mere effektivt at selektere direkte for overlevelse. På den anden side er der enhøjere arvbarhed for fødselsvægt i forhold til for overlevelse, hvilket betyder, at det godtkan være gavnligt at inddrage fødselsvægt som en ekstra egenskab sammen med avl foroverlevelse (33). Selvom den genetiske korrelation mellem fødselsvægt og overlevelse errelativt lav gør den højere arvbarhed af fødselsvægt, at det vil kunne have væsentligeeffekter på overlevelse, hvis begge egenskaber inddrages i et selektionsindeks (33). Manbør være opmærksom på, om effekten af fødselsvægt er lineær, eller der er en ikke lineærsammenhæng (f.eks. 25, 26). Roehe et al. (2009) anbefaler at selektion skal være for en”optimal” fødselsvægt for at undgå en øget dødelighed i forbindelse med faringen af tungegrise (33). Knol et al. (2002) konkluderer i overensstemmelse hermed, at selektion for øgetfødselsvægt vil kunne resultere i flere dødfødte grise (78).Udvikling af nye, og mere effektive selektionsmetoder gennem genomisk selektion, vil pålidt længere sigt desuden medføre, at især egenskaber med lav arvbarhed, så somoverlevelse, vil kunne inddrages mere effektivt i avlsarbejdet. I et igangværende projektom genomisk selektion undersøges effekten af at inddrage overlevelse i avlsarbejdet hosøer som får mulighed for at passe 14 levende grise. I projektet ses på derfor påoverlevelse i kuld der er standardiseret til 14 grise efter fødsel. Baggrunden for valg afnetop dette mål for overlevelse er et kombineret ønske om at selektere for øgetoverlevelse og selektere for søer, der kan passe og fravænne et stort kuld. Forventningener, at dette projekt vil kunne danne basis for at inddrage egenskaber i krydsningsgrisen iavlsarbejdet, herunder overlevelse. Dette arbejde udføres som et samarbejdsprojektmellem Videncenter for Svineproduktion og Århus Universitet og forventes at løbe indtil2015. Genomisk selektion er allerede under implementering i Duroc.26
PATTEGRISDØDELIGHED I DK
Som det fremgår af ovenstående er det muligt allerede nu at inddrage overlevelse somselektionskriterium i det danske avlsprogram. Tidshorisonten for en effekt af inddragelse afoverlevelse i avlsarbejdet vil afhænge af, hvornår og hvordan egenskaben vægtes iavlsindekset. For de orner der udvælges i dag, vil de første af deres krydsningssøstre fødegrise om ca. 20-22 måneder. Den fulde effekt af denne selektion kommer derefter løbendeefterhånden som ornernes døtre kommer i brug i produktionsbesætningerne i de følgendeår. Hertil skal lægges tid til at implementere nye tiltag i registreringssystemer, databaserog avlsværdivurdering. Nu og her kan vægtningen af LG5 i avlsmålet ændres. Detteforventes dog ikke at være et tiltag der vil påvirke overlevelsen væsentligt, dadødeligheden er stagneret efter indførslen af LG5 i avlsmålet. Det vil dog reducerefremgangen i kuldstørrelse og dermed mindske stigningen i antal døde per kuld.Rentabilitet i soproduktionen skal imidlertid sikres gennem en optimering af antalfravænnede grise og den indsats, der skal lægges i at få grisene levende frem tilfravænning. Derfor vil der være en økonomisk balance mellem om det fortsat kan betalesig at øge antallet af levende grise gennem avlen, og den pris det har, at man samtidigskal sikre en bedre overlevelse gennem bedre (og sandsynligvis dyrere) management. Deter her vigtig at bemærke, at det har en økonomisk pris at øge vægten på overlevelse iavlsindekset. Et stort antal levende grise i kuldet vil altid kræve en bedre management endfå levende grise i kuldet, uanset avlskriterium. Så længe antallet af levende grise forventeshøjere end antallet af funktionsdygtige patter kan man tillige forvente, at indsatsen afmanagement skal øges uproportionalt mere, da denne situation stiller højere og andrekrav til so og grise. Derfor er det fortsat vigtig at have fokus på nye management tiltag,som kan sikre bedre overlevelse.
3.1.2
Fødsel af større og mere levedygtige grise gennem fodring afsoen sidst i drægtighed
Adskillige forsøg er blevet udført med henblik på at undersøge effekten af den drægtigesos foderoptagelse på fødselsvægt. Mange af disse undersøgelser har haft fokus påkødkvaliteten og har derfor set på effekten af øget foderoptagelse i perioden fra ca. dag 25til dag 80 – 90 i drægtigheden, hvor muskelfibrene dannes. Generelt påvirker en øgetfoderoptagelse ikke fødselsvægten, og kun ekstreme reduktioner i proteinindtag harindflydelse på afkommets fødselsvægt (79).Folinsyre og B12 vitamin er sat i forbindelse med antallet af dødfødte grise (80).Vitaminerne er nødvendige for syntese af DNA og RNA, hvilket vil påvirke de stamceller,som danner de røde blodlegemer. Ved mangel på vitamin B12 og folinsyre vilproduktionen af røde blodlegemer falde, hvilket vil medføre en reduktion i blodetsilttransportkapacitet og dermed indebære risiko for underforsyning af fostrene med ilt.Antallet af levendefødte grise synes at kunne øges ved tildeling af meget høje doser affolinsyre til soen omkring løbning eller i den første del af drægtigheden. Dette ændrer dogikke umiddelbart det grundlæggende problem med, at en stor del af pattegrisedødelighedskyldes grise, som fødes med lav fødselsvægt. Udenlandske forsøg har vist en stigning iantallet af levendefødte grise ved en tildeling af folinsyre til drægtighedsfoderet på op til27
PATTEGRISDØDELIGHED I DK
950 % af den danske norm og ved injektion af 15-750 mg folinsyre 2 til 10 gange påforskellige tidspunkter omkring løbning og i den første halvdel af drægtigheden (81-83).Matte et al. (1984) fandt imidlertid ikke en forøgelse af kuldvægten ved fødsel, og der varen tendens til en øget variation i kuldvægt indenfor kuld (83). Effekten på den samledepattegrisedødelighed er derfor vanskelig at vurdere. På baggrund af forsøg med rottersynes effekten af folinsyre at være ledsaget af en øget mængde af RNA, DNA og protein ifostrene (84), og jo højere RNA koncentration i fosteret, desto større eroverlevelseschancen. Folinsyre synes også at virke indirekte ved at stimulere sekretionenaf vækstfaktorer i uterus samt faktorer af betydning for implanteringen af fostret i uterussom f.eks. prostaglandin E2(85).Der findes kun begrænset viden om B12 vitamins indflydelse på reproduktionen. En nydansk afprøvning af effekten af injektion af B12 vitamin til drægtige søer viste hverkeneffekt på total kuldstørrelse, antal dødfødte grise eller forekomsten af farefeber (80).Tilsætning af L-carnitin til den drægtige sos foder kan derimod være en mulighed for atøge fødselsvægten. L-carnitin dannes naturligt i kroppen ud fra aminosyren lysin.L-carnitins primære rolle er at transportere lange- og mellemkædede fedtsyrer overmitokondriernes indre membran som substrat forβ-oxidationi fedtforbrændingen.Desuden er L-carnitin involveret i protein syntese og glukose homeostase. Indflydelsen afL-carnitin tilsætning til drægtige søer har i tre ud af 4 studier vist, at fødselsvægten varøget med ca. 100 g, når den drægtige so blev fodret med 100 til 125 mg/dag af L-carnitin(79).Tilsætning af aminosyren, L-arginin, er også en mulighed. L-arginin er en naturligtforekommende aminosyre. I en undersøgelse blev L-arginin tilsat (0 eller 1 %) foderet tildrægtige gylte fra d 30 til faring. Dette forsøg viste, at det totale antal fødte pattegrise varupåvirket, hvorimod antallet af levende fødte pattegrise steg fra 9,37 til 11,40 vedtilsætning af 1 % L-arginin til foderet (86). I dette forsøg indgik 52 gylte. Derudover findesder resultater fra andre dyrearter og mennesker, der støtter, at L-arginin har betydning forfostervækst. Således er det fundet, at fodring med 50 % af normen til drægtige fårresulterer i nedsat fødselsvægt. Tilsætning af L-arginin ophæver denne negative effekt afreduceret foderoptagelse på afkommets fødselsvægt (35). Ligeledes mindskes IUGR hosmennesket efter behandling med L-argining (87). Virkningen af L-arginin synes at kunnetilskrives en øget blodforsyning til fostrene og en stimulering af fostrenes vækst ogudvikling. Blodgennemstrømningen i placenta er bl.a. reguleret af nitrogen oxid (NO), somstimulerer dannelse af blodkar og forårsager karudvidelse. Dette reaktive stof dannes udfra aminosyren arginin og katalyseres af enzymet nitrogen oxid syntase (NOS). Endvideredannes polyaminer udfra arginin, og disse stimulerer celledeling og udvikling af celler tilspecifikke væv (35). Tilsætning af L-arginin til foderet i drægtigheds- ogdiegivningsperioden i gylte er ligeledes vist at øge pattegrisenes vækst idiegivningsperioden specielt fra dag 0 til dag 7 (88). Dette var en konsekvens af, at defleste aminosyrer i mælken var øget efter fodring med L-arginin.
28
PATTEGRISDØDELIGHED I DK
En anden vigtig faktor for grisenes medfødte mulighed for overlevelse er, at deresglykogendepoter ved fødslen. Da grise til forskel fra f.eks. mennesker fødes med en megetbegrænset mængde omsættelige fedtdepoter, er glycogendepoterne stort set de enesteenergireserver grisene har, og størrelsen af disse depoter er derfor essentielt for grisenesmulighed for at overleve eventuel energimangel eller kulde (Figur 8 – 89, 90). Det er vist,at grisenes medfødte glykogendepoter kan forøges ved tilsætning af mellem-kædedefedtsyrer til soens foder fra 2-4 uger før faringen, og at dette øger grisenes overlevelse(91). Tilsætning af mellem-kædede fedtsyrer til soens foder den sidste uge før faringkunne derimod ikke påvirke glykogendepoterne (92).Også faringsforløbet synes at kunne påvirkes af ernæringsmæssige faktorer, især dekomponenter, der indvirker på dannelsen af prostaglandin F2α, som er den naturligeigangsætter af faringen og også anvendes ved faringsinduktion. Sådanne komponenterkan bl.a. være zink (93) og linolsyre (94). Byggestenen for prostaglandin F2αerarakidonsyre, der dannes ud fra n-6 fedtsyren linolsyre, hvis tilgængelighed afhænger afforholdet mellem n-3 og n-6 fedtsyrer samt E-vitamin status (95). Zink formodes at havebetydning for den enzymatiske omdannelse på flere trin i omdannelsen af linolsyre tilprostaglandiner (96).Det er påvist, at en lav zinkstatus kan medføre forsinket og forlænget faring hos søer (97),mens øget zinktildeling reducerer faringens varighed (98) og øger grisenes overlevelse,bl.a. gennem en reduceret andel af dødfødte grise og en forbedret overlevelse aflevendefødte grise (98-100). Årsagssammenhængene er ikke velkendte, men tilskrivesmanglende uterus sammentrækninger forårsaget af ovennævnte nedsatte syntese afprostaglandiner. Svins labile zinkpulje er meget lille fra naturens hånd (98) og kan derforkan blive utilstrækkelig i fysiologisk kritiske perioder som ved faring, især hvis søernesfodertildeling begrænses i perioden op til faring.Betydningen af foderets fedtsyresammensætning og E-vitamin indhold omkring faring forfaringsforløbet er ikke belyst, men det er veldokumenteret, at både E-vitamin indholdet ogfedtsyresammenætningen i soens blod og mælk kan påvirkes af fodringen (101,102). Etøget fokus på mineral- og vitaminbehovet i relation til foderets indhold af n-6 og n-3fedtsyrer omkring faring og viden om prostaglandin dannelsen med henblik på at reducerefaringskomplikationer vil være relevant i fremtidig forskning og fastsættelse affodringsmæssige anbefalinger.
3.1.3
Fødsel af større grise og mere levedygtige grise gennemforbedret management
Management i drægtigheden og omkring faring har betydning både for grisenesfødselsvægt og for farings forløb. Flere undersøgelser har vist, at stress i drægtighedenmedfører et fald eller en tendens til et fald i fødselsvægt (103-107), mens andre ikke harkunnet dokumentere denne effekt (108-112). En enkelt undersøgelse viste endog forøgetfødselsvægt hos grise, der var udsat for simuleret stress i den sidste del af drægtigheden(113). De varierende resultater afspejler formodentlig, at det er afgørende, hvornår idrægtigheden belastningen finder sted, og at stress kan have uheldige følger for29
PATTEGRISDØDELIGHED I DK
fødselsvægten. I undersøgelser af betydningen af socialt stress inden dag 105 idrægtigheden, der afspejler en af de typiske belastninger for danske søer, sås ikkenegative effekter på hverken fødselsvægt eller pattegrisedødelighed (110, 112).Stress i den sidste del af drægtighed synes derimod at øge risikoen for faringsproblemer.Faringsforløbet reguleres via oxytocin og nedsat frigivelse af oxytocin er forbundet medforlængede faringer (114, 115). Effekten af stress på faringsforløbet går sandsynligvis viaen inhibering fra stress hormoner på frigivelsen af oxytocin, som medvirker til at regulerefødselsforløbet (116) Faringsproblemer i form af lange fødselsintervaller øger den tid,grisene opholder sig i fødselskanalen og udgør en risiko for, at grisene dør på grund afiltmangel under fødslen (31, 48, 117). Forlænget ophold i fødselskanalen svækker ogsåde levendefødte grise og øger derfor risikoen for, at de senere dør af andre årsager, somf.eks. sygdom, dårlig trivsel o. lign. (31).To typiske kilder til stress i forbindelse med faringen er sen indsættelse og/eller fiksering afsoen i forbindelse med faring. Det er vist i flere undersøgelser at fiksering i farestaldenmedfører forhøjet niveau af stress hormonet cortisol (115, 118, 119) og øgethjertefrekvens (120). Samtidig er der påvist reduceret koncentrationer af oxytocinsammenfaldende med forlængede faringer (115). Førstegangsfiksering af unge søer, derflyttes i kassestier, har i flere andre undersøgelser ud over en negativ effekt påfaringsforløbet også resulteret i en øget andel af dødfødte grise, både ved rettidigindsættelse (13, 121) og ved sen indsættelse i kassestien (122). Flere undersøgelser harogså fundet forlænget faring hos ældre fikserede søer sammenlignet med søer indsat ikassestier (49, 123). Der blev dog ikke fundet nogen forskel i antallet af dødfødte grise pået mindre antal søer mellem søer indsat i kassestier og løsdriftsstier som alle blev indsat igod tid og fik tildelt store mængder af redemateriale (124), eller som blev flyttet fra løsdrifttil enten en fareboks med fiksering eller en løsdriftssti i god tid (7 dage) før forventet faring(21). Effekten slår sandsynligvis stærkest igennem på dødelighed, når unge søerførstegangsfikseres og under forhold hvor introduktionen til farestien er suboptimal ogbidrager til at øge belastningen (f.eks. sen flytning og manglende halm).Risikoen for faringsproblemer på grund af førstegangsfiksering af gylte må forventes atstige ved fuld implementering af løsdrift i drægtighedsstalden i alle besætninger fra 2013,idet langt den overvejende del af besætningerne fortsat vil fiksere søer og gylte ifarestalden. Hvis dette tillige kombineres med sen indsættelse, hvilket er en typisk følge afunderdimensionering af antallet af farestipladser, vil der være risiko for, at gylte belastes ien sådan grad, at det kan øge andelen af dødfødte grise pga. faringskomplikationer. Dagyltekuld normalt udgør omkring 25 % af alle kuld i farestalden, kan konsekvenserne bliveanselige. Andelen af gyltekuld er først og fremmest styret af niveauet for udsætning afsøer med dårlige produktionsegenskaber, og økonomiske beregninger tyder på, atudskiftningsprocenten med fordel kan reduceres (125). Også gennem udvælgelse af mererobuste gylte eller ved reduktion af sodødeligheden kan andelen påvirkes. Tilvænning tilfarestien for gyltene kan lettes ved at anvende løsdrift i hele soens reproduktive cyklus, ogved at fremtidssikre dimensioneringen af staldene. Fremtidssikring af staldene vilindebære, at der dimensioneres med et antal stipladser i farestalden, der sikrer at alle søer30
PATTEGRISDØDELIGHED I DK
og gylte kan flyttes til farestalden mellem 7 og 5 dage før forventet faring, samtidig med atder afsættes stipladser til ammesøerne, der med den nuværende kuldstørrelse er ennødvendig del af produktionen. Det er her vigtigt at pointere, at behovet for ekstrastipladser til ammesøer yderligere vil stige, hvis der gennem forskellige tiltag opnås denønskede reduktion i dødelighed.Sygdomme hos soen, så som klovlæsioner (126) og blærebetændelse (127) har vist sig atøge antallet af dødfødte grise. Effekten kan være en følge af, at sygdomme genereltmedfører en stressreaktion hos soen. Sygdomme hos soen vil derfor muligvis generelthave en negativ effekt på antallet af dødfødte grise. En forbedring af sundhedsstatus forsøerne vil sandsynligvis kunne reducere andelen af dødfødte grise. Tillige har en enkeltundersøgelse vist, at der er højere risiko for dødfødte grise, når rumtemperaturen er over22 �C i forhold til under 22 �C (128).Udover disse problemstillinger har flere undersøgelser vist, at antallet af dødfødte grisekan reduceres markant gennem overvågning og udøvelse af faringshjælp ved behov.Undersøgelserne har vist, at intensiveret faringsovervågning og hjælp til nyfødte pattegrisekan halvere den totale dødelighed både hos fikserede (42, 129, 130) og løsgående søer(131). I en gennemsnitlig dansk besætning vil det betyde, at den totalepattegrisedødelighed sænkes fra 24 % til 12 % af kuldet. Bemærkelsesværdigt viserundersøgelserne også, at andelen af grise, der betegnes som ”dødfødte” (typisk 5-10 % afkuldet) rent faktisk kan overleve ved rettidig indgriben i faringen eller lige efter grisenesfødsel. Det er desuden vist, at udførelse af faringshjælp, når fødselsintervallet overstiger 1time, kan reducere andelen af søer med farefeber (132). De nødvendige tiltag er imidlertidarbejdskrævende, især da fødslen af hver enkelt gris skal overvåges manuelt, såindgrebene kan gøres rettidigt. Holdfaringer med store hold, eventuelt samlet over 3 ugersom ved 3-ugers drift, vil i højere grad muliggøre, at det bliver økonomisk rentabelt at laveen sådan faringsovervågning. Det vil desuden være hensigtsmæssigt at udvikleautomatiserede metoder, der kan overvåge faringsforløbet og varsle landmanden vedlange fødsler. Dette kan være en hjælp til at målrette arbejdsindsatsen mod de kuld, dervirkelig har faringsproblemer og lade kuld uden problemer i fred. Indledendemetodeudvikling til detektion af fødselsintervaller baseret på billedbehandlingsmetoder fravideooptagelser er netop igangsat ved DJF men kræver yderligere forskning og udvikling,før de kan implementeres.Faringsinduktion ved injektion af prostaglandin F2αanvendes i nogle besætninger sommiddel til at synkronisere faringen. Dette kan være medvirkende til, at faringsovervågningkan praktiseres (133). Nogle undersøgelser har vist, at faringsinduktion kan reducereforekomsten af farefeber, andre at det øger risikoen for farefeber (134), mens andre igenhar vist en reduceret råmælksproduktion ved faringsinduktion (135). En ældre danskbesætningsundersøgelse kunne imidlertid ikke påvise, at faringsinduktion medprostaglandin mellem dag 112 og 115 i drægtigheden påvirkede antal levende- ellerdødfødte eller pattegrisedødeligheden i diegivningsperioden (136).
31
PATTEGRISDØDELIGHED I DK
3.2
Sikring af at grisen er i stand til hurtigt at komme tilpatterne og indtage råmælk
Som omtalt i kapitel 2 afhænger grisenes overlevelsesevne af, at de er i stand til hurtigt atkomme hen til yveret og få råmælk. En af de vigtigste forudsætninger for dette er en størrefødselsvægt og vitalitet, der kan forbedres gennem ovennævnte tiltag. Hertil er detafgørende, at underafkøling kan undgås, både gennem klimamæssige tiltag, tildeling afhalm og valg af hensigtsmæssige gulve.En række undersøgelser har vist, at underafkøling af de nyfødte pattegrise i deresallerførste levetimer har negativ betydning for grisenes overlevelse både i kassestier (21,36, 57), løsdriftsstier (21, 56) og udendørssystemer (50). I systemer med meget halm,f.eks. udendørssystemer, hvor soen kan bygge en rede, kan halmen hjælpe med til atbevare varmen omkring soen, så grisene fødes i et optimalt klima (137). I danskebesætninger anvendes sjældent halm i farestien og kun i meget små mængder. Dertildeles typisk ekstra varme til grisene i et hjørne af stien, men alle undersøgelser viser, at idet første levedøgn, hvor behovet for varme er ekstra stort, anvender grisene ikke enopvarmet hule i nævneværdig grad (138-140). Der er derfor behov for andre måder atsikre et optimalt klima omkring de nyfødte pattegrise. Det er vist, at varme i gulvet påfaringsstedet i løsdriftsstier signifikant øgede pattegrises kropstemperatur i de førstelevetimer, reducerede tiden til første råmælksoptagelse og forbedrede pattegrisenesoverlevelse (61). Tilførsel af varme på faringsstedet kræver, at stien er indrettet med enstor andel fast gulv, da det er vanskeligt at tildele varme i et spaltegulv. Alternativt kanindlæggelse af varmebevarende måtter (141) sandsynligvis give en lignende effekt. IDanmark opstaldes i dag næsten alle faresøer i kassestier med helt eller delvistspaltegulv, og der vil typisk være en stor andel spaltegulv på fødselsstedet og ved soensyver. Der er derfor behov for at videreudvikle og teste metoder til tilførsel af varme til denyfødte pattegrise på anden måde, end ved simpel opsætning af varmelampe, somtidligere har været afprøvet (142). DJF er p.t. i gang med at teste og dokumentere virkningaf en alternativ varmekilde til nyfødte pattegrise hos fikserede søer på grisenes evne til atopretholde normal kropstemperatur i de første levetimer efter fødsel. På længere sigt kandet være hensigtsmæssigt at udvikle nye farestikoncepter, som kan håndtere størremængder af halm således, at de varme- og optørrende egenskaber ved halm kan udnyttestil at reducere nyfødte pattegrises varmetab og derved sikre overlevelse.
3.3
Sikring af grisens mulighed for at indtage råmælk/mælk
Råmælken udgør grisenes primære kilde til energi og antistof i de første leveuger og eressentiel for grisenes overlevelse. Ved faringens start vil der være en vis mængde råmælki soens patter. Efter de nyfødte grise har fjernet den første råmælk, vil mælken i defølgende timer udskilles i perioder på 1-4 minutter med intervaller på mellem 5-30 minutter.Omkring 6-12 timer efter faringens start begynder mælkenedlægning gradvist at ændrekarakter. Mælken nedlægges nu i kortere og kortere perioder, og der bliver længeremellem diegivningerne. I løbet af det første døgn er diegivningerne helt etableret.
32
PATTEGRISDØDELIGHED I DK
Mælkenedlægningen varer nu kun ca. 20 sek., og der vil som regel være ca. 45-60 minmellem hver diegivning. Samtidig med at diegivningerne ændrer karakter, begyndergrisene at danne en patteorden, og deres dieadfærd bliver synkroniseret. Grisene vil nu såvidt muligt die fra én og samme patte ved hver diegivning, og de vil forsvare denne patteoverfor andre grise i kuldet. Diegivningen foregår efter et fast mønster, som er etkomplekst samspil mellem so og grise. Grisenes evne til at fjerne mælk fra den enkeltepatte, og deres ihærdighed med at massere patten, påvirker mælkeydelsen i den enkeltepatte. Det vil sige, at soens mælkeproduktion og fordeling af mælk til de enkelte patter ikkealene er bestemt af soens arveanlæg og næringsstofforsyning, men også af den enkeltegris’ evne til at stimulere patten (143) og forsvare den overfor kuldsøskende (144, 145).Soen har typisk 14-16 funktionelle patter i den første laktation, og antallet falder medstigende laktationsnummer. Grundet det specielle diegivningsmønster, hvor grisene harderes faste patteorden, og mælken kun er til rådighed for grisene i et ganske kort intervalca. én gang i timen, kan en so maksimalt give die til et antal grise, der svarer til denspatteantal.Det er afgørende for grisenes mulighed for at overleve, at de hurtigt finder en patte ogigennem de første levedage bliver i stand til at etablere ejerskab til den. I kuld med fleregrise, end soen har patter til, vil det være nødvendigt at flytte overskydende grise tilammesøer, hvis de skal kunne overleve. Hvis brug af ammesøer søges undgået, er detderfor nødvendigt at tilpasse avlen for antal grise i forhold til avl for patteantal.
3.3.1 Betydning af stimiljø for pattegrise adgang til og stimulation afpatterneFor at sikre en optimal optagelse af råmælk/mælk, er det vigtigt, at pladsforholdene ifarestien tillader, at alle pattegrise kan die uforstyrret. I en kassesti kan adgangen tilpatterne være begrænset af inventaret. Mange kassestier har begrænset plads, når soenhviler på den ene side, idet stierne er usymmetriske (figur 7). Ved fødslen vil der dog somudgangspunkt være tilstrækkelig plads i de fleste stityper til, at grisene kan ligge vinkelretud fra soens yver. Derimod kan deres adgang vanskeliggøres af bøjler, placeret lodreteller vandret ud fra soens yver. F.eks. er det vist at variationen i tilvækst blev forøget vedbrug af horisontelle rør i stedet for vertikale rør (146). Desuden giver de fleste kassestier ideres nuværende form ikke tilstrækkelig plads til, at grisene kan die uhindret, når de bliverstørre (se Bilag 1). Mange kassestier er ikke tilstrækkeligt brede til, at grisene kan ligge ideres fulde længde og die på begge sider af soen. Fire uger gamle pattegrise måler igennemsnit 56 cm i længden (147). At pladsforholdene påvirker pattegrisenes tilvækst, erpåvist i flere undersøgelser både i kassestier (148) og i løsdriftsstier (149). Andreindretningsmæssige forhold der har betydning for grisenes stimulation af soens patter oghendes efterfølgende mælkeproduktion er støjniveauet i rummet. Som det fremgår Af Bilag1 er der et væsentligt støjniveau i mange farestalde. Undersøgelser har vist at støjforringer kommunikationen mellem pattegrise og so under mælkenedlægningen (150). Støjførte til at pattegrisene brugte kortere tid på at masserer patterne hvorved soensmælkeproduktion blev reduceret (143).
33
PATTEGRISDØDELIGHED I DK
Figur 7. Kassestier er almindeligvis usymmetriske. Foto: M.K. Bonde
3.3.2 Brug af ammesøerEn anden vigtig faktor for pattegrisenes mulighed for at få næring nok i form afråmælk/mælk er kuldstørrelsen. Grise som fødes i store kuld og som fødes sidst ifødselsrækkefølgen har f.eks. optaget mindre antistof som følge af mindreråmælksoptagelse end grise som fødes først og/eller fødes i små kuld (151). Af hensyn tiloptagelsen af råmælk/mælk er det derfor vigtigt at der ikke er flere grise end der er pattetil. Søernes råmælksydelse synes ikke at afhænge af kuldstørrelsen (152). Mængden afråmælk, der er til rådig for den enkelte gris, er derfor ofte mindre i store kuld og antallet afdiende grise har derfor også betydning for både dødelighed og tilvækst (153).Soen har som omtalt typisk 14-16 funktionsdygtige patter, og i mange kuld fødes der i dagmere end 20 levende grise. Råmælksforsyningen til disse pattegrise vil jf. ovenståendevære reduceret i forhold til hvis de fødes i mindre kuld, hvilket betyder nedsatimmunforsvar og større risiko for sygdom. Kuldudjævning og brug af ammesøer er medden stigende kuldstørrelse trods dette et nødvendigt redskab til at give alle grise adgang tilen funktionel patte og redde overskydende grise fra at dø (154). Kuldudjævning foretagesså vidt muligt efter at grisene har optaget råmælk men indenfor det første levedøgn. Detgøres ved at overskydende pattegrise flyttes fra kuld med mange grise (typisk mere end12 levende) til kuld med færre grise. På den måde udjævnes kuldstørrelsen ved fødsel.Kuldudjævning, specielt hvis den foretages senere end det første levedøgn, har negativekonsekvenser for diegivningsadfærden i form af afbrudt diegivning i op til 6 timer (155,156), ustabilitet i patteorden (156-159), aggression fra soen rettet mod fremmede grise(155-158), og at nye grise ikke umiddelbart bliver integreret i kuldet og ligger mere påspaltegulvet og udenfor hulen (156). Dertil kommer, at tilvæksten typisk er forringet hos
34
PATTEGRISDØDELIGHED I DK
tilflyttede grise (157, 159-162). Hos soens egne grise i kuld med kuldudjævning har manogså fundet et fald i tilvækst sammenlignet med ikke-kuldudjævnede kuld (157), om enddette ikke var så stort som for de tilførte grises. Pattegrisedødeligheden er ofte højere forde nytilkomne grise (154) og er større ved sen end tidlig udjævning (163). Tidspunktet forflytning har også indflydelse på tilvæksten, idet den negative effekt på tilvækst er størreved sen flytning (164).I mange besætninger foretages kuldudjævning tillige med det formål at få så ensartedekuld som muligt rent vægtmæssigt. Begrundelsen for denne procedure synes at være athindre store grise i at dominere mindre grise, så de ikke får adgang til patterne. Ingennyere undersøgelser bekræfter imidlertid, at der kan opnås fordele i form af højere tilvæksthos alle grise ved at sortere dem efter størrelse ved fødsel (165, 166). Derimod er fundeten reduktion i tilvækst på omkring 20 % (163).Den succesfulde selektion for øget kuldstørrelse i de seneste år har medført, at antallet aflevende grise ved fødslen i dag i gennemsnit er på 14,2 grise i Danmark. Indtil for få årsiden kunne en overproduktion af pattegrise håndteres ved hjælp af traditionelkuldudjævning, mens de fleste landmænd i dag rutinemæssigt anvender ammesøer ogbudgetterer med stipladser til disse. Brug af en ammeso adskiller sig fra traditionelkuldudjævning ved, at soen modtager et helt nyt kuld grise, der som regel er sammensataf overskudsgrise fra andre kuld, og ikke kun et antal grise som supplement til sit egetkuld. En ammeso giver først die til egne grise i typisk 3 uger og giver dernæst die tiloverskydende grise fra nyfødte kuld, der typisk er få dage gamle. Det har været anbefalet,at lade søer die max. 12 grise, og en ny undersøgelse bekræfter, at det har konsekvenserfor tilvækst og overlevelse, hvis soen har 15 grise frem for 11 eller 13 grise ved yveret(153). Så ud fra en antagelse om, at 12 grise fortsat er almindelig praksis, kan detberegnes, at med det nuværende antal levende grise ved fødsel, er der behov for, at ca.15 % af alle søer også indgår som ammesøer. Der er dog store forskelle i, hvordan dettehåndteres i praksis, og generelt mangles dokumentation for konsekvenserne af deforskellige angrebsvinkler. Alternativet til at bruge kuldudjævning og ammesøer vil være, atet stort antal af pattegrisene må aflives for ikke at dø af sult eller tilhørende årsager, hvilketikke er etisk og velfærdsmæssigt forsvarligt.Velfærd hos ammesøerDer er ikke foretaget egentlige undersøgelser af ammesøers velfærd. Når en so bliveranvendt som ammeso, betyder det, at dens diegivningsperiode forlænges til 6-7 uger istedet for ca. 4 uger som er den gennemsnitlige diegivningsperiode i Danmark. Alt andetlige betyder det, at soen skal fikseres i en længere periode med mange velfærdsmæssigekonsekvenser til følge (32). Desuden er der undersøgelser, som tyder på, at en langdiegivning under produktionsforhold, hvor soen ikke kan forlade grisene, kan værebelastende. Disse undersøgelser er dog ikke entydige.Søer vil gennem diegivningsforløbet gradvist forsøge at vanskeliggøre grisenes adgang tilyveret for at sikre grisene en gradvis overgang til fast føde. Under forhold hvor soen ikke
35
PATTEGRISDØDELIGHED I DK
kan vælge at fjerne sig fra pattegrisene eller undgå deres pattestimulering (f.eks. itraditionelle kassestier), kan det derfor muligvis være belastende for ammesoen at skulleopretholde en diegivning i 6-7 uger. Det er fundet, at søer i farebokse prøver at hindrepattegrisenes adgang til yveret ved at ligge mere på maven 4 uger efter faring end tidligerei diegivningsperioden (167, 168). Ligeledes er det vist, at søer reducerer frekvensen afdiegivninger igennem laktationsperioden, hvis de har mulighed for det (169, 170).Fravænningsprocessen er gradvis og starter allerede fra 2. leveuge men varer typisk fra10-16 uger. Tendensen til at soen reducerer grisenes adgang til yveret er påvirket afsoens huld og kuldets størrelse, således at søer i godt huld og med små kuld er meretilbøjelige til at give die end søer med store kuld og stort vægttab (171). Det er derforsandsynligt, at ammesøer i godt huld og med få fostergrise i mindre grad vil værebelastede af den længere diegivning end søer i dårligt huld og med mange fostergrise.Der er kun få undersøgelser, hvor der er foretaget målinger af stresshormon (frit cortisol,ACTH) hos søer gennem diegivningen (172, 173). I undersøgelserne blev det basaleniveau af cortisol målt i blodet frem til dag 28, men resultaterne er modstridende. I den eneundersøgelse blev der fundet en stigning omkring dag 28, som var mere udtalt hosfikserede søer end hos løse søer (172), hvilket antyder, at fiksering kombineret med enlængere diegivning kan være belastende. I den anden fandtes der ingen stigning, hverkeni cortisol eller ACTH, og det havde ingen betydning, om søerne var fikserede eller løse(173).Konsekvenser af en lang diegivningsperiode for soens vægt er heller ikke entydige. Der erundersøgelser, der viser, at søernes vægttab ikke var påvirket af diegivningsperiodenslængde (174, 175). En anden undersøgelse viste derimod, at hvis søerne kan gå væk frapattegrisene og har færre diegivninger, så taber de sig mindre (176). En undersøgelsetyder på, at løsgående søer i forhold til fikserede søer er bedre i stand til at undgåvægttab, når grise efterspørger mere mælk, idet de fandt højere foderforbrug og størretilvækst hos pattegrisene, når søerne gik løse i forhold til, når de var fikserede. Derimodvar der ingen forskelle i vægttab for søerne (149). En forøget risiko for skuldersår hosammesøer kunne umiddelbart forventes, men der er ikke påvist øget forekomst afskuldersår hos ammesøer (177).
3.3.3 Avl der sikrer, at kuldstørrelse er tilpasset antal patterFlere undersøgelser har vist at antallet af patter er arvelig (bl.a. 162). Der er dog ikkefundet evidens for, at avl for øget patteantal resulterer i en reduceret dødelighed. Antalpatter kunne dog være én af flere maternelle egenskaber, der kunne bidrage til en øgetoverlevelse. Antal patter og andre maternelle egenskaber relateret til pattegriseoverlevelse kunne være indikator for egenskaber, der bidrager til en mere sikkerudvælgelse af genetisk overlegne søer for pattegrise overlevelse, men de er ikke i sig selvavlsmål. Dette forudsætter dog, at deres genetiske sammenhæng med overlevelse bliverundersøgt.
36
PATTEGRISDØDELIGHED I DK
3.4
Søer der producerer tilstrækkeligt råmælk/mælk af højkvalitet
Det har afgørende betydning for pattegrisenes vitalitet og overlevelse, at soens mælk, isærråmælken, produceres i en mængde og energimæssig kvalitet, der sikrer grisenesenergibehov lige efter fødslen. Det skyldes, at grisenes medfødte energidepoter kun kandække grisenes behov i ca. 16 timer, og at depoterne ikke må udtømmes indenproduktionen af overgangsmælk etableres (Figur 8 – 89, 90).Energiforsyning til nyfødte grise100
80råmælkovergangs mælk60so mælkGlykogen depot40
20
0012345
Dage efterfødsel
Figur 8: Figuren viser, hvorledes den nyfødte gris forsynes med energi. I det første levedøgn er bådeoptagelsen af råmælk og de medfødte glykogendepoter livsvigtige for at grisen kan overleve på kort sigt.Kilde: Theil et al., 2010 (90).
Råmælkens immunologiske kvalitet er også vigtig, da den skal sikre grisene en passivimmunitet overfor infektiøse sygdomme og fremme etablering af en mikroflora i tarmen,der både beskytter mod opblomstring af sygdomsfremkaldende mikroorganismer ogbidrager til udnyttelse af mælkens antistoffer (f.eks. 179-181).Uanset det potentiale soen har for en tilstrækkelig råmælksproduktion af god kvalitet, erdet helt afgørende, at soen ikke får farefeber, idet dette almindeligvis medførermælkemangel (182,192).Både mængden og kvaliteten af råmælken er uafhængig af kuldstørrelsen og kan derforvære en begrænsende faktor for grisenes overlevelse i store kuld, især for gyltekuld, hvorbåde mængde og kvalitet af råmælken er ringere og for kuld fra ældre søer, hvormængden af råmælk er lavere (f.eks. 152, 183, 184). Betydningen i praksis afproduktionen af råmælk i forhold til grisenes evne til at sikre sig adgang til råmælken erdog ikke veldokumenteret, men grisenes overlevelsesevne øges med optagelsen afråmælk (Figur 9).37
PATTEGRISDØDELIGHED I DK
0-24 timers overlevelse (%)
1009896949290400450500550600Optag af råmælk (g pr gris pr dag)
y = 0.048x + 73r = 0.84
Figur 9. Sammenhæng mellem råmælksoptagelse og pattegrisenes overlevelse i første levedøgn. Kilde:Theil, 2010 (89).
3.4.1 Energiindhold og mængde af råmælkDen energimæssige kvalitet af råmælk og tidlig mælk afhænger i høj grad af enhensigtsmæssig fodring i sendrægtigheden, da denne både påvirker mængden ogkvaliteten af råmælken (152) og risikoen for, at søerne får mælkemangel på grund affarefeber (182, 192). På Videncenter for Svineproduktions hjemmeside (www.vsp.lf.dk)findes opdaterede anbefalinger vedrørende fodersammensætning og foderstrategi idrægtighed og diegivningsperioden, der på baggrund af forskning og afprøvning bedstmuligt tilgodeser mælkeproduktionen under samtidig hensyn til reproduktions- ogproduktionsegenskaberne på længere sigt. Fodringen af danske søer tager dog kun ibegrænset omfang højde for, at produktionen af råmælk og mælk stimuleres forskelligt.Ligeledes er normerne for næringsstoftildeling til søerne ikke generelt justeret i forhold tilden store kuldstørrelse.Nyere forskningsresultater peger på, at de fodringsmæssige anbefalinger de sidste 4 ugeraf drægtigheden kan forbedres og derved øge grisenes overlevelse. Tilsætning af mellem-kædede fedtsyrer til soens foder er i flere undersøgelser fundet at forbedre pattegrisenesoverlevelse (91, 185-187). I nogle af undersøgelserne kan effekten til dels have skyldtesen forbedret opbygning af grisenes glykogendepoter, idet tildelingen skete fra 2-4 uger førfaringen (91, 185), mens den i undersøgelser, hvor tildelingen kun er sket i den sidste ugeinden faring, hvor glykogendepoterne ikke længere kan påvirkes (92), skyldes forbedretindhold af energi i mælken (186, 187).Tilsætning af hydroxy methyl butyrate eller mellem-kædede fedtsyrer til sofoderet de sidste3-7 dage før faring er vist at kunne øge fedtindholdet i råmælken (186, 187) og den tidlige38
PATTEGRISDØDELIGHED I DK
mælk (188). Desuden forøger tildeling af fedtkilder med mange mellem-kædede fedtsyrerden tidlige mælks indhold af mellem-kædede fedtsyrer, som grisene kan udnytte på ligefod med kort-kædede fedtsyrer indenfor det første levedøgn (188, 189). Ioverensstemmelse hermed er der vist reduceret dødelighed i det første levedøgn (fra 5 %til 0 %) ved tildeling af oktansyre til foderet, dog på et lille datamateriale (8 søer perbehandling) (187). Flere naturlige foderemner, som f.eks. kokosolie, indeholder storemængder af mellem-kædede fedtsyrer, men der er behov for yderligere klarlægning afvirkningsmekanismer og forsyningskilder, før det kan implementeres i praksis.
3.4.2 Immunologisk kvalitet af råmælkMed hensyn til den immunologiske kvalitet af råmælken har der overvejende været fokuspå overførsel af antistoffer og især på overførsel af specifikke antistoffer modtabsvoldende sygdomme hos grisene. Sidstnævnte tilgodeses i dag ved vaccination af dekommende moderdyr mod de mest tabsvoldende pattegrisesygdomme og udsættelse afde kommende moderdyr for små mængder af den besætningsspecifikke smitte (190, 191).I forhold til de gældende vejledninger vedrørende vaccination vil flytning af andenvaccination, der normalt gives 2-3 uger før faring, til 6 uger før forventet faringsandsynligvis forbedre råmælkens indhold af antistoffer mod den specifikke vaccine (193).Dette tiltag kan være relevant på besætningsniveau. Da pattegrisedødelighedenforårsaget af de specifikke infektioner, der typisk vaccineres imod, i dag synes at være lav(22, 194), er det dog ikke sandsynligt, at et sådant tiltag i sig selv vil påvirkepattegrisedødeligheden nævneværdigt på nationalt plan. Om muligt vil udvikling af eneffektiv vaccine mod ny spædgrise diarré, der optræder med stigende hyppighed i landemed intensiv svineproduktion (22, 194-197), derimod kunne have et potentiale for atreducere dødeligheden på nationalt plan. Dette forudsætter imidlertid øget kendskab tildenne diarrés ætiologi, hvilket søges opnået i et igangværende samarbejdsprojekt mellemKøbenhavns Universitet, DTU og Videncenter for svineproduktion (198).Vaccination øger ikke råmælkens totale indhold af antistoffer, men øger antistoffer modden specifikke vaccine (152). Den totale mængde antistoffer synes at kunne øges vedtildeling af produkter baseret på uspecifikke immunstimulatorer (bl.a. hydroxy methylbutyrate), gær (Saccharomycescerevisiae),fermenteret vådfoder eller probiotika (135,199). Virkningsmekanismerne er ikke kendte, men det er foreslået, at effekten er knyttet tilmodulerende virkninger på mave-tarm kanalens mikroflora, idet der er et tæt samspilmellem det lokale immunsystem i tarmen og dannelse af antistoffer til råmælken (135,200). Desuden kan antistofindholdet øges ved tildeling af særlige fedtsyrer, somkonjugeret linolsyre og visse olier (135, 199).I nogle undersøgelser var det forøgede antistof indhold i råmælken ledsaget af enreduceret pattegrisedødelighed ved tildeling af gærprodukt, immunstimulatorer, hydroxymethyl butyrate eller probiotika (199, 201, 202). Produkterne påvirkede imidlertid ogsåandre egenskaber, der kan forklare eller medvirke til en forbedret overlevelse. En danskundersøgelse fra 1997 kunne ikke påvise en effekt af probiotika påpattegrisedødeligheden (203). I betragtning af de seneste års ændring i avlsmaterialet vil
39
PATTEGRISDØDELIGHED I DK
det dog være relevant at undersøge dette igen. Det er uvist, om den totale mængdeantistoffer i råmælken indenfor normalområdet er afgørende for grisenes overlevelse,forudsat at grisene får den råmælk, der er nødvendig ud fra en energimæssig betragtning,og at betingelserne for optagelse af antistofferne over tarmen i form af råmælkens indholdaf hjælpestoffer og et optimalt termisk miljø for grisene er til stede. Da små grise har enforbedret evne til at optage antistoffer over tarmen (204), er råmælkens totaleantistofindhold i sig selv næppe begrænsende for grisenes overlevelse i store kuld, hvis deblot får tilstrækkelig råmælk.Den immunologiske kvalitet af råmælken er dog meget mere end et spørgsmål omantistoffer. Råmælken indeholder også stoffer, der fremmer grisenes evne til at optageantistofferne over tarmen, hvor bl.a. vitamin A og E kan øges gennem soens fodring idrægtigheden (102, 180). Desuden indeholder råmælken faktorer, der modner tarmen,beskytter mod opblomstring eller invasion af patogener eller er immunsupprimerende ogderved muliggør etablering af en naturlig mikroflora (179, 180, 205-207). Endelig overføresder med råmælken immunceller, som har immunmodulerende effekter på grisen (152, 206,208, 209). Råmælkens indhold af selen, vitamin A, C og E, og særlige fedtsyrer menes athave betydning for grisenes immunkompetence gennem indflydelsen på den anti-oxidativekapacitet. Af disse vitaminer og mineraler er det kun C vitamin, som ikke kan påvirkesgennem fodring af soen (152). Råmælkens antistofindhold og øvrige immunologiskekvalitet kan givet optimeres gennem fodring, men flere af de processer, som sikresgennem råmælkens kvalitet, er modsatrettede. Derfor er der behov for yderligereforskning, hvor der ses på den samlede effekt af manipulationer og betydningen forgrisenes overlevelse, før dette kan udnyttes i praksis.
3.4.3 Forebyggelse af farefeberFarefeber eller MMA omfatter yverbetændelse (mastitis), børbetændelse (metritis), og/ellermælkemangel (hypo- eller agalactia). De kliniske symptomer er nedstemthed,appetitløshed, feber, forstoppelse, yverbetændelse, flåd fra vulva og mælkemangel (182,192).De bakterier, der hyppigst er isoleret fra mælkekirtler fra søer med MMA erE.coli,Staphylococcus aureusogepidermis,hæmolytiske Streptococcer ogKlebsiella spp.,oftestKlebsiella pneumonia,men disse bakterier findes også i mælkekirtler fra raske søer (182,192). Dette tyder på, at der er tale om sekundære infektioner med opportunistiske,potentielt patogene bakterier (192). Risikoen for farefeber øges derfor typisk i situationer,hvor immunforsvaret er svækket.En dansk undersøgelse viste en gennemsnitlig forekomst af farefeber på 9,5 % i periodenfra 1976-1982 (192). Der findes ikke nyere landsdækkende opgørelser, men antallet afsøer, der behandles mod farefeber, angives at være ca. 20 % (182). Forekomsten erformentlig lavere, da en vis overbehandling må forventes at finde sted. Det kan dog ikkeudelukkes, at der er sket en stigning i forekomsten, da risikoen for farefeber øges medkuldstørrelsen (192, 127), især ved sen indsættelse i farestalden (134). Sen indsættelsesker formodentlig med stigende hyppighed som følge af det øgede behov for stipladser til40
PATTEGRISDØDELIGHED I DK
ammesøer. Desuden kan ændringen i opstaldningsform mod øget hyppighed af skift fraløsdrift i drægtigheden til boksopstaldning i farestalden (se 1.3) øge forekomsten affarefeber (210-212). Sygdomskomplekset kan medføre en ekstra pattegrisedødelighed på6-9 % (192).Farefeber menes ofte at være knyttet til fedme og forstoppelse hos soen sandsynligvisfordi både fedme og forstoppelse medfører forlængede faringer (49, 209). Det forebyggesgennem hensigtsmæssig fodring i henhold til anbefalingerne, huldstyring, sikring af en godvandforsyning, et godt klima, god hygiejne og ved at minimere belastning af soen i densidste tid før faringen (182, 192), og ved at give soen bevægelsesfrihed omkring faring(49).Risikoen for farefeber vil muligvis kunne reduceres yderligere gennem tiltag, derstabiliserer søernes tarmflora og modvirker forstoppelse (omtalt under 3.4.2) om enddanske afprøvninger vedrørende dette ikke har vist positive effekter i forhold til farefeber(203). Set i lyset af den generelt forøgede risiko for farefeber, ovennævnte potentielleeffekter på råmælkens kvalitet (3.4.2) og den nyeste viden om betydningen af den normaletarmflora for immunsystemets funktion, vil det dog være relevant at undersøge effekten afprobiotika nærmere.
3.4.4 Håndtering af farefeberVed farefeber skal soen tidligst muligt behandles efter de retningslinjer, der foreligger(182). Det er vigtigt for grisenes overlevelse at sikre, at grisene får råmælk og energi. Hvissoen ikke er i stand til at give denne anbefales det, at give grisene råmælk fra andre søer ibesætningen eller eventuelt mælkeerstatning. Desuden anbefales det, at give griseneadgang til vand (182). Muligheden for at kunne gribe ind rettidigt forbedres vedovervågning af faringerne (3.1.2).
3.5
Påpasselige søer og kvikke grise
Risikoen for ihjellægning afhænger både af soens påpasselighed over for grisene og afgrisenes evne til at flytte sig, når soen ruller eller lægger sig. Der er stor forskel på deenkelte søers påpasselighed overfor grisene. Søernes moderegenskaber er i et vistomfang medfødte men afhænger også af deres senere erfaringer. Herudover påvirkersoens sundhed dens påpasselighed overfor grisene. Tilsvarende afhænger grisenes evnetil at flytte sig af, at de ikke bliver syge. I en australsk undersøgelse var 44 % afihjellægningerne knyttet til sygdom hos enten so eller grise (213).
3.5.1 Udvælgelse af søer med gode moderegenskaberDer er foretaget en del undersøgelser af sammenhæng mellem søernesmoderegenskaber og pattegrisedødelighed. Flere undersøgelser tyder på, at søer, somhar stor aktivitet under redebygningen (31, 214) og ligger mere stille efter faringsstart
41
PATTEGRISDØDELIGHED I DK
(215), har færre ihjellægninger. En enkelt undersøgelse på få søer antydede tillige, at søermed kraftig reaktion på griseskrig (214) havde færre ihjellagte grise. Ligeledes på et lilledatamateriale er det vist, at søer med kraftig reaktion på griseskrig, og som samtidighavde få positurskift, lagde færre grise ihjel (216). Flere forfattere har desuden vist, atsøer, der forberedte grisene på forestående lægge-sig adfærd og først lagde sig, når derikke var grise i nærheden (217) havde færre ihjellægninger. Endelig er det vist i flereundersøgelser, at søer som var mere forsigtige, når de lagde sig, også havde færreihjellagte grise (214, 217-219).Da der samtidig synes at være en vis gentagelighed fra kuld til kuld i soens moderadfærd(220-222), er der foretaget undersøgelser af, om det var muligt at udvælge gode mødreenten via avlen eller blot via udvælgelse af polte med en adfærd, som var korreleret tilgode moderegenskaber. Der er fundet koblinger mellem poltes generelle reaktivitet,frygtsomhed overfor mennesker og stressreaktioner og den efterfølgende moderadfærd(221-223).Et forsøg på at vise denne kobling på et større materiale ved brug af kortvarige og lethåndterbare tests under produktionsrelevante forhold viste dog, at sammenhængen varforholdsvis svag (215), hvilket gør, at der p.t. ikke findes nogen dokumenteret måde atudvælge polte med forventede gode moderegenskaber i produktionssammenhæng.Selektion for en kraftig reaktion i tests med afspilning af griseskrig og håndtering af grise,lav frygtreaktion overfor mennesker, og lavt aggressionsniveau i forbindelse meddannelsen af nye sociale grupper synes derimod at resultere i kuld med lavere dødelighed(224-226). En anden metode til vurdering af søers moderegenskaber uden et storttidsforbrug er anvendelsen af subjektive vurderinger af søernes reaktion på enstandardiseret test eller en vurdering af soens adfærd over en længere periode (225, 227).Generelt findes dog en lav arvbarhed for adfærdsmæssige moderegenskaber og en lav tilmoderat genetisk korrelation mellem adfærdsmæssige moderegenskaber ogpattegrisedødelighed, hvilket formodentlig skyldes, at miljøet og de aktuelle forholdomkring de enkelte kuld spiller en stor rolle i forhold til de genetiske egenskaber ved so oggrise. Generelt er det problematisk i denne type undersøgelser, at der kuldudjævnes, såsoen ikke udelukkende passer sit eget afkom, og at der er en dårlig diagnostik afdødsårsager, som oftest baseret på en visuel inspektion, hvilket betyder stor usikkerhedom grisenes reelle dødsårsåger. Inddragelse af maternelle adfærdsegenskaber iavlsarbejdet alene for at reducere pattegrisedødelighed vil derfor have en meget lilleeffekt, og det vil være mere hensigtsmæssigt at lave en direkte avl for bedre overlevelse(3.1.1).
3.5.2 Stimulering af soens moderegenskaber ved tildeling af plads oghalmDer findes god dokumentation for, at søernes påpasselighed over for grisene øges ogrisikoen for ihjellægning nedsættes, hvis søer får mulighed for at bygge rede. Mangeundersøgelser har vist, at søer er stærkt motiverede for redebygning op til faring (f.eks. 32,228). Samtidig kan adgang til halm stimulere redebygningsaktiviteten (f.eks. 229-231). Det
42
PATTEGRISDØDELIGHED I DK
er påvist, at høj aktivitet under redebygning (31, 214, 232) og lav aktivitet under faringen erforbundet med en lavere risiko for ihjellægning (215). Efter faring blev antallet afpositurskift hos både fikserede søer (233) og løsgående søer (234) reduceret vedhalmtildeling, og antallet af nærklemninger blev reduceret hos løsgående søer, som fik friadgang til hel halm (229, 235). Tillige er det vist, at fikserede søer med meget halm somunderlag og overdækning af boksen viste en højere reaktivitet overfor grise i nød (236).Hos løsgående søer er det påvist, at feedback fra en rede resulterede i, at de nyfødtegrise hurtigere fandt patterne og fik adgang til råmælk (237). Undersøgelse af hvilkenmængde hel halm søer anvendte dagligt i dagene før, under og efter redebygning iløsdriftssystemer viste, at søer i dagene før og efter faring i gennemsnit anvendte ca. 0,5kg hel halm dagligt. På redebygningsdagen anvendte søerne derimod i gennemsnit ca. 1,5kg hel halm med en variation fra <0,5 kg til 7,5 kg (238).To undersøgelser på et større antal søer har set på effekten af at tildele ekstra halm påpattegrisedødelighed. Den ene er lavet på fikserede søer, hvor søerne fik tildelt 100 gversus 1000 g snittet halm til redebygning (239). Den anden er lavet på løsgående søer,hvor søerne fik tildelt 2,5 kg hel halm sammen med 4 kg snittet halm eller 4 kg snittet halmuden hel halm (240). Ingen af undersøgelserne fandt en effekt på pattegrisedødelighed.Søers mulighed for at udvise moderadfærd kan tillige stimuleres ved at give søernebevægelsesfrihed, f.eks. i form af løsdrift eller udendørssystemer. Det er kendt at løsdriftbåde reducerer risikoen for at soens bider nyfødte pattegrise ihjel (121, 241), detøger/ændrer redebygningsaktiviteten (228, 242), øger søers reaktivitet overfor skrig fraklemte grise (222) og øger søernes mulighed for at orientere sig om hvor grisene befindersig før de lægger sig, hvilket har betydning for risikoen for ihjellægning (242). Til trods forat løsdrift stimulerer moderadfærd af betydning for ihjellægning er der dog ikke fundetlavere ihjellægning i systemer hvor søerne går løse. I flere undersøgelser, hvor dødeligheder sammenlignet mellem løsgående søer og fikserede søer, er der derimod fundet, at fleregrise dør af ihjellægning i løsdriftsstier, mens flere grise dør af andre årsager i systemerhvor søerne fikseres (243, 244). Sandsynligvis afspejler disse forskelle primært, at deundervægtige og underudviklede pattegrise dør under alle omstændigheder, men at degør det af lidt forskellige årsager. Hos løsgående søer er der større mulighed for, at soenlægger sig på en svag gris, som er ved at dø, mens en sådan svag gris i en kassesti oftevil ligge og dø i et hjørne af stien, hvor soen ikke har adgang pga. fikseringen. Samtidig errisikoen for, at en løsgående so lægger sig på en allerede død gris også større. Grisefundet døde under soen vil blive registreret som værende ihjellagt, selvom dette ikkenødvendigvis er den udløsende årsag. En anden forklaring kan også være, at de nævnteundersøgelser alle er baseret på landmandens egen vurdering af dødsårsagen og ikke påbasis af obduktionsfund. Resultatet kan derfor også afspejle landmandens forventning til,hvad årsagen til dødelighed er i de forskellige systemer, hvor de fleste vil være meretilbøjelige til at registrere en lille gris som værende klemt i et løsdriftsystem end i enkassesti.
43
PATTEGRISDØDELIGHED I DK
3.5.3 Forebyggelse af sygdom hos soenSygdomme, der medfører febertilstande, lidelser i bevægeapparatet og muligvis ogsåsmertefulde lidelser hos den farende og diegivende so synes at øge risikoen forihjellægning (213, 192, 245-247). Smertelindring af søer med farefeber ved anvendelse afde såkaldte NSAID midler kan føre til en reduktion af pattegrisedødeligheden, især forgrise med lav fødselsvægt (248), men den produktionsøkonomiske effekt ved at anvendemidlerne i danske undersøgelser har været begrænset (182). Efter de gældende regler erbrug af smertelindrende NSAID midler begrænset til anvendelse ved farefeber ogledbetændelse samt i undtagelsestilfælde, hvor dyrlægen skønner det nødvendigt (249).Ud fra den generelle hyppighed af forskellige lidelser hos danske søer vil en intensiveretforebyggelse af farefeber, ben- og klovlidelser, skuldersår og mave-tarm lidelsersandsynligvis kunne reducere pattegrisedødeligheden på nationalt plan. Forebyggelsespecifikt af farefeber er omtalt tidligere.Forekomst og karakter af sygdomme hos søerne er besætningsspecifik, og behovet for ogkarakteren af forebyggende tiltag og effekten af disse vil derfor også værebesætningsafhængig. Et igangværende projekt hos Videncenter for Svineproduktion harvist, at sodødeligheden på besætningsniveau kan reduceres væsentlig ved atimplementere kendt viden gennem en intensiveret rådgivning, især med fokus på huld,ben- og klovlidelser, samt gulve og sygestier. De foreløbige resultater viser, atsodødeligeheden kunne reduceres i 14 ud af de 17 besætninger, der indgik i projektet(250, 251). Nedbringelsen af sodødeligheden medførte dog kun i én ud af tre besætningeren lille stigning i antal fravænnede grise (251). Anvendelse af erfaringerne fra projektet tilen fortsat intensiveret rådgivning omkring sygdomsforebyggelse, især lidelser ibevægeapparatet, og en eventuel smertelindring må forventes at kunne reducere den delaf pattegrisedødeligheden, der hidrører fra ihjellægning. En reduktion af sygdomme idrægtighedsperioden synes tillige at kunne reducere andelen af dødfødte grise (3.1.3).Videncenter for svineproduktion har for nylig på deres hjemmeside annonceret, at de haren målsætning om, at projektet, Soliv, skal nedbringe sodødeligheden med en fjerdedelinden 2013.På længere sigt vil intensiveret sygdomsovervågning ved hjælp af IT-baseret automatiskregistrering af sygdomsmarkører formodentlig kunne øge muligheden for rettidige indgrebmod sygdomme. De første initiativer til udvikling af et sådant redskab er iværksat iDanmark.Der er os bekendt ikke kendskab til, hvordan stigningen i kuldstørrelse påvirker søernesholdbarhed og velfærd. Det er dog ikke utænkeligt, at det stigende pres på søerne i formaf større kuldstørrelse og øgede krav til stor mælkeydelse udgør en belastning for soen,som kan reducere dens holdbarhed. Undersøgelser af disse sammenhænge bør belysesnærmere ved fortsat avl for øget kuldstørrelse.
44
PATTEGRISDØDELIGHED I DK
3.5.4 Reduktion af sygdom hos griseneSom for sygdomme hos soen er hyppigheden og karakteren af sygdomme hos grisenebesætningsspecifik. På nationalt plan skønnes sygdomme hos grisen at være ansvarlig forca. 22 % af pattegrisedødeligheden. Diarré (enteritis) og generaliseret infektion(blodforgiftning og ledbetændelse – septicæmi/arthritis) er de hyppigste årsager tilinfektiøst betinget pattegrisedød (Figur 6 - 21).Som omtalt i pkt. 3.4 vil tiltag, der forbedrer grisenes immunitet ved at øge råmælkensimmunologiske kvalitet eller reducere forekomsten af farefeber, reducere risikoen forsygdom hos grisene. Desuden forebygges sygdomme hos grisene gennem minimering afsmittepresset i farestalden. Dette omfatter tiltag som alt-ind-alt-ud drift i farestalden,højtryksrensning, desinfektion og tørring af farestalden før indsættelse, spaltegulv i en delaf farestien, minimering af kuldudjævning, hensigtsmæssige arbejdsrutiner påbesætningsniveau og eventuel vask af søerne i mild sæbe og vand før indsættelse ifarestalden.Desuden er der en væsentlig risiko for spredning af infektioner gennem sår og rifter pågrisene. Skader på grisene er typisk klovskader og læsioner på forknæ samt læsioner iansigtet som følge af pattekampe. Risikofaktorer for skader på klove, ben og forknæ ergulvbelægningen. Et blødt underlag i form af f.eks. neoprenmadrasser (141), spagnum(252) eller en større mængde halm (253) reducerer risikoen for skader på ben og dervedrisikoen for halthed og efterfølgende aflivning eller generaliserede infektioner. Ruoverflader er ligeledes en risikofaktor, f.eks. ældre beton gulve (252) eller specielleskridsikre underlag (254), mens andre har fundet øget risiko for læsioner ved spaltegulveaf stål frem for plastic og jern (255).En øget specifik indsats mod pattegrisediarréerne er vanskeliggjort af den øgedeforekomst af en ny type spædgrise-diarré, hvis årsag ikke kendes (22). De generaliseredeinfektioner vil muligvis kunne nedbringes ved optimering af gulvene i farestier, så gulvenepå én gang tager hensyn til en høj stihygiejne og en lav risiko for læsioner på grisenesknæ, idet sådanne kan udgøre en indgangsport for smitte. Dette område har alleredefokus i svineproduktionen, da farestaldens gulve også har betydning for forekomsten afskuldersår.
3.6
Sammenfatning
Sammenfattende er der god dokumentation for, at pattegriseoverlevelsen kan øges ved atinkludere en økonomisk vægtning af dødelighed evt. i samspil med fødselsvægt iavlsindekset. Der er også potentielle muligheder for at øge pattegrises fødselsvægtgennem optimeret fodring af de drægtige søer, men der foreligger p.t. ikke tilstrækkeligdokumentation på området, ligesom der heller ikke er tilstrækkelig dokumentation forgennem fodring at forbedre faringsforløbet og reducere antal dødfødte grise gennemfodring. Det synes vigtigt, at reducere stress under drægtigheden og i særdeles at undgå45
PATTEGRISDØDELIGHED I DK
stress i forbindelse med flytning til farestalden, f.eks. sen indsættelse og skift fra løsdrift tilfiksering. Faringsovervågning har potentiale for at reducere både andelen af dødfødtegrise og andelen af grise, der dør efter faring. Mange grise dør af underafkøling, og sikringaf et bedre klima på fødselsstedet gennem tildeling af ekstra varme, isolerende underlageller rigelig med strøelse kan reducere dødeligheden. Fødsel af kuld med flere grise, endsoen har patter betyder, at grisenes adgang til råmælk og mælk begrænses. Derfor er detnødvendigt at foretage kuldudjævning og bruge ammesøer. Disse tiltag kan hindreoverskydende grise i at dø, men de pågældende grise vil dog have reduceret tilvækst ogøget risiko for at dø i forhold til deres kuldsøskende, som forbliver ved egen mor. Denprimære velfærdsmæssige konsekvens af brug af ammesøer er den øgede tid søerne skalvære fikserede og muligvis, at de ikke kan udøve kontrol over diegivningerne, når de erfikserede.Både mængden og kvaliteten af råmælken kan være en begrænsende faktor for grisenesoverlevelse i store kuld. Både den energimæssige og den immunologiske kvalitet afråmælken synes at kunne forbedres gennem fodring. Der er dog behov for yderligereforskning, før det kan implementeres i praksis. Uanset det potentiale soen har for entilstrækkelig råmælksproduktion med god kvalitet, er det helt afgørende, at soen ikke fårfarefeber, da dette almindeligvis medfører mælkemangel.Risikoen for ihjellægning afhænger både af soens påpasselighed over for grisene og afgrisenes evne til at flytte sig, når soen ruller eller lægger sig. Der er stor forskel på deenkelte søers påpasselighed overfor grisene, men inddragelse af maternelleadfærdsegenskaber i avlsarbejdet vil have en meget lille effekt i forhold til direkte avl forbedre overlevelse. Soens påpasselighed over for grisene kan øges, hvis søer fårbevægelsesfrihed og mulighed for at bygge rede, og hvis sygdomme, der medførerfebertilstande, lidelser i bevægeapparatet og muligvis også smerte, kan undgås.Tilsvarende afhænger grisenes evne til at flytte sig af, at de ikke bliver syge. Soens oggrisenes sundhed tilgodeses i dag bedst ved god hygiejne, faringsovervågning,hensigtsmæssige gulve og klima, tiltag, der forbedrer grisenes immunitet, og vedintensiveret rådgivning med fokus på soens sygdomme.
46
PATTEGRISDØDELIGHED I DK
Kapitel 4
Potentielle tiltag, der kanreducere pattegrisedødeligheden
Som det fremgår af afsnit 3 findes der kun få tiltag, som umiddelbart vil kunne nedbringepattegrisedødeligheden i Danmark.Inddragelse af overlevelsesraten direkte i selektionskriteriet i det danske avlsprogram vilvære en oplagt mulighed indenfor et kortere tidsrum. Tidshorisonten for en effekt afinddragelse af overlevelse i avlsindekset vil afhænge af, hvordan egenskaben vægtes iavlsindekset, samt hvor hurtig avlsarbejdet slår igennem i produktionsbesætningerne. Pålængere sigt vil der muligvis kunne ske en yderligere optimering af avlsarbejdet i forhold tilgriseoverlevelse f.eks. gennem genomisk selektion eller inddragelse af overlevelse ihandyrracen Duroc. Det forberedende arbejde til denne effektivisering er allerede igangsataf erhvervet. Det vil yderligere være muligt at forbedre avlen for overlevelse ved atinddrage én af flere maternelle egenskaber, som f.eks. antallet af patter eller pattegrisenesfødselsvægt, som indikator egenskaber. Der er således et behov for også fremover atforbedre de genetiske modeller, så de bedre beskriver kuldstørrelse ogdødelighed/overlevelse, og at undersøge de genetiske sammenhænge til øvrigeegenskaber i avlsmålet.En intensiv indsats på managementområdet, både i form af intensiveretfaringsovervågning fulgt op af rådgivning med henblik på reduktion af sygdomme hossoen, skønnes principielt at kunne medføre op til en halvering af pattegrisedødelighedenmed virkning indenfor en kort tidshorisont. Faringsovervågning, hvor soen ydesfaringshjælp efter behov og det sikres, at grisene ikke bliver underafkølet, og at de fåradgang til råmælk/mælk, vil kunne redde mange grise både blandt de, der ellers dør underfaringen og blandt de små, svage grise, der dør af sult eller kulde eller senere erhverversygdom pga. et svagt immunforsvar. Faringsovervågning er imidlertid arbejdskrævende,da fødslen af hver enkelt gris skal overvåges manuelt, så indgrebene kan gøres rettidigt.Hvis der skal være økonomisk dækning for faringsovervågningen, vil det sandsynligviskræve, at antallet af søer, der farer samtidig, optimeres i forhold til den arbejdstid, dermedgår til overvågning. På grund af den naturlige variation i løbningstidspunkt og idrægtighedslængde vil faringerne kunne ske over de fleste af ugens dage. Dog vil omkring60-70 % af søerne fare indenfor 2-3 dage. En økonomisk rentabel faringsovervågning afalle søer vil derfor primært kunne ske i store besætninger, eller i mindre besætninger, hvorsøernes faringer er samlet i faringshold f.eks. hver 3. uge frem for hver uge.Faringsinduktion synkroniserer i et vist omfang faringerne, og givet på det rigtige tidspunktaf dagen vil behandlingen kunne føre til, at 70 procent af søerne vil fare i dagtimerne, hvorfaringsovervågningen er mindst omkostningstung. Faringsinduktion med prostaglandinmedfører dog i sig selv omkostninger, da den skal foretages af en dyrlæge, tidligst mågøres 3 døgn før forventet faring og på grund af variationen i løbningstidspunktet måforventes at medføre 2 dyrlægebesøg. Desuden vil faringsinduktion betyde, at en visprocentdel af søerne farer 2-3 dage før rettidig faring, hvilket kan have negative
47
PATTEGRISDØDELIGHED I DK
konsekvenser for grisenes vitalitet ved fødsel. De nævnte begrænsninger betyder, atpotentialet i en intensiveret faringsovervågning sandsynligvis ikke kan udnyttes fuldt ud pånationalt niveau, uden at det har en økonomisk omkostning.Projektet, Soliv, der sigter mod nedbringelse af sodødeligheden og løber indtil 2013,indebærer samtidig en intensiveret rådgivning med henblik på reduktion af sygdomme hossøer (250). Hvis dette implementeres i alle problembesætninger med fokus på sygdommei farestalden vil det potentielt kunne påvirke pattegrisedødeligheden positivt, idetrådgivningen øger fokus på områder, der har betydning for pattegrisedødeligheden, somf.eks. polteudvælgelsen (3.1.3). En lavere sodødelighed vil resultere i en lavere andel afgylte i besætningen (3.1.3) og være relateret til en øget sundhed hos søerne med en deraffølgende reduktion i alle typer af pattegrisedødelighed (3.1.3, 3.4.3 og 3.5.3). Ud fra deresultater, der er offentliggjort fra projektet, Soliv (251), ser metoden lovende ud for atnedbringe sodødelighed, men foreløbig med en begrænset effekt på pattegrisedødelighedefter faring. En sådan rådgivning er imidlertid også økonomisk omkostningstung og skalderfor holdes op mod, hvad der kan opnås ved f.eks. intensiveret overvågning.På sigt vil omkostninger i form af arbejdslønninger til både intensiveret faringsovervågningog sundhedsstyring på besætningsniveau kunne reduceres i det omfang, der udviklesautomatiserede metoder til overvågning af faringsforløbet og varsling af landmanden vedlange fødsler samt til sygdomsovervågning af soen. De indledende trin i en sådanudvikling er påbegyndt, men yderligere forskning og udvikling er nødvendig, førautomatiseret overvågning kan implementeres. Der er således behov for fortsat fokus pådenne type forskning.I en del besætninger vil pattegrisedødeligheden sandsynligvis også i nogen grad kunnereduceres gennem et optimeret stimiljø med forbedrede termiske forhold for grisene, og enreduceret infektionsrisiko. Sådanne tiltag vil f.eks. være alternative varmekilder til nyfødtepattegrise (3.2.1) og forbedrede gulve (3.5.4).Ændring til løsdrift i farestalden kan have et potentiale for at reducere antallet af dødfødtegrise ved at belastning af soen på grund af skift mellem opstaldningssystemer minimeres.Tiltaget vil sandsynligvis primært slå igennem på de 25 % af faringerne, som ergyltefaringer. Alternativt kan det tillige have en effekt på antallet af dødfødte grise ogsandsynligvis også på døde efter faring, at der ved nybyggeri og gennem tilpasning afbesætningsstørrelsen sikres farestipladser nok, så alle søer kan indsættes minimum 5-7dage før forventet faring.På lidt længere sigt vil der formentlig også gennem intensiveret forskning kunne udviklesredskaber i form af ernæringsmæssige tiltag i drægtigheden, der kan reducerepattegrisedødeligheden. Flere specifikke komponenter synes enten at øge grisenesoverlevelsesevne samlet set eller at øge grisenes fødselsvægt (3.1.2), deresglykogendepoter (3.1.2), reducere risikoen for faringskomplikationer (3.1.2), øgefedtindholdet i råmælken og den tidlige mælk (3.4.1), alle faktorer, der vil bidrage til størreoverlevelse hos pattegrisene. Potentielle fokusområder er L-carnitin, L-arginin, zink, E-
48
PATTEGRISDØDELIGHED I DK
vitamin, og fedtsyresammensætning – især mellem-kædede fedtsyrer. Desuden kanprobiotika være interessant, men den tilgængelige dokumentation på området er megetbegrænset. Generelt er der behov for yderligere dokumentation af virkningerne, ikkemindst under produktionsforhold, hvor mange andre faktorer også påvirkerpattegrisedødeligheden. Desuden vil der være behov for afklaring af forsyningskilder ogforsyningssikkerhed, før det kan implementeres i praksis.Råmælkens kvalitet, i form af antistoffer og bioaktive komponenter, der bidrager tiludvikling af funktionen af grisenes mavetarmkanal, kan givet også optimeres gennemfodring, men betydningen af dette for grisenes overlevelse kendes endnu ikke (3.4.2).Flere af de processer, som sikres gennem råmælkens kvalitet, er modsatrettede, og der erbehov for en betydelig forskningsindsats, hvor der ses på den samlede effekt afmanipulationer og betydningen for grisenes overlevelse, før en forsvarlig indsats kaneffektueres i praksis. Desuden er der behov for øget kendskab til den nye spædgrisediarrés ætiologi, så der om muligt kan udvikles effektive vacciner mod denne.Hvis de foreslåede management tiltag iværksættes i danske besætninger, kan detforventes, at antallet af diende grise i kuldet stiger. En fortsat avl for LG5 vil desuden øgeantallet af grise i kuldet yderligere. Selv med indførelse af et evt. nyt avlsmål, sominkluderer overlevelse, vil der stadig være en mulighed for at antallet af grise, som dier,forøges, afhængig af hvor stor økonomisk vægt, der sættes på dødelighed i indekset. Dade eksisterende farestier allerede nu ikke synes tilstrækkelig store til at kunne rumme soog pattegrise, jf. opmåling af danske søer (10, 147) og kassestier (bilag 1), er detnødvendigt, at der ved nybyggeri investeres i større kassestier og/eller løsdriftsstier. Etstigende antal levende grise i kuldet vil tillige betyde et stigende behov for brug afammesøer. De velfærdsmæssige konsekvenser ved at en so bliver til ammeso ser primærtud til at være i form af længere tids fiksering og evt. via dens manglende mulighed for atreducere grisenes adgang til yveret og derved at udøve en vis kontrol over egenmælkeydelse. Ved at holde ammesøer i løsdrift med et begrænset antal ammegrise vil develfærdsmæssige konsekvenser sandsynligvis kunne reduceres. Det er dog muligt, at deter vanskeligere at få ammesøer i løsdrift til at acceptere de fremmede grise, da enløsgående so i højere grad kan reagere mod de fremmede grise.
49
PATTEGRISDØDELIGHED I DK
Kapitel 5
Konklusion
Danske søer har igennem avlen opnået en af verdens højeste kuldstørrelser, menDanmark er også det land, som har den højeste andel døde grise ud af det totale antalfødte grise sammenlignet med de lande, vi har fundet oplysninger fra. Dette skyldes enandel af grise, der enten er dødfødte eller dør indenfor de første timer efter faring.Opgørelserne fra besætninger giver ikke mulighed for at vurdere, hvor meget der kantilskrives et øget antal dødfødte grise, og hvor meget der skyldes dødsfald efter fødsel,men litteraturgennemgangen tyder på, at dødelighed i begge kategorier øges medkuldstørrelsen. Den høje andel af døde grise er derfor både et etisk og velfærdsmæssigtproblem, da de fleste grise, som dør efter fødslen, synes at gøre det ved en lidelsesfulddød.Som det fremgår af afsnit 1 har de seneste års avl mod større kuldstørrelse formodentligværet en stor del af årsagen til den stigende andel af døde grise i perioden fra 1992 til2004. Inddragelse af antal levende grise ved dag 5 som avlsmål i stedet for kuldstørrelsesynes ikke at være et avlsmål, der kan nedbringe andelen af døde grise, men målet harført til en stagnering af den totale dødelighed af pattegrise.Den store kuldstørrelse har nødvendiggjort brug af ammesøer for at hindre overskydendegrise i at dø ved deres biologiske mor som følge af manglende patter til alle grise. Brug afammesøer udgør primært et velfærdsmæssigt problem, fordi det betyder, at ammesøernefikseres i en længere periode. Desuden medfører brug af ammesøer et stigende behov forfarestipladser, som bør imødekommes for at sikre, at søerne kan indsættes i farestalden igod tid før faring.Indførelse af pattegriseoverlevelse i det danske avlsindeks er en oplagt mulighed for atreducere pattegrisedødelighed, og de nødvendige oplysninger og beregningsmetoder erallerede tilgængelige eller kan fremskaffes med begrænset indsats. Dette vil dog ikke føretil en reduktion her og nu af pattegrisedødelighed i produktionsbesætningerne.Tidshorisonten for en effekt af inddragelse vil afhænge både af, hvordan overlevelsevægtes i avlsindekset, og hvornår avlsarbejdet slår igennem i produktionsbesætninger(første effekter kommer tidligst efter 20-22 mdr.). Der er desuden forskningsaktiviteter igang, som på sigt kan bidrage til at effektivisere avlen for egenskaber, som f.eks.overlevelse, vækst og patteantal, yderligere.Der vil fortsat være behov for en intensiv indsats på managementområdet for at reducerepattegrisedødeligheden, da den høje kuldstørrelse stiller store krav til so og management.Der er god dokumentation for, at forbedret management i form af øget faringsovervågningog intensiveret rådgivning omkring sosundhed nu og her vil kunne forbedre grisenesoverlevelse. Et bedre nærklima f.eks. i form af halm eller opsætning af ekstra varmekilder,hvor pattegrisene fødes vil hindre, at nyfødte pattegrise underafkøles straks efter fødslen.Dette vil sandsynligvis kunne reducere dødeligheden. Desuden viser flere undersøgelser,at en risikofaktor for dødfødte grise er det skift fra løsdrift til fiksering, som søerne
50
PATTEGRISDØDELIGHED I DK
udsættes for, når de flyttes til farestalden. Dette problem øges med fuld implementering afløsdrift i alle drægtighedsstalde fra 2013. Indførelse af løsdrift i hele reproduktionscyklusvil kunne afhjælpe dette problem og samtidig sikre bedre velfærd for søerne, men kan ikkeforventes overordnet at føre til en reduktion i pattegrisedødeligheden. På sigt vil flereandre tiltag kunne vise sig at have en positiv indflydelse, men disse forudsætter yderligereforskning, udvikling eller afprøvning.
Referencer1.Videncenter for Svineproduktion. Årsberetninger.http://vsp.lf.dk/Publikationer/Aarsberetninger%20VSP.aspx2.Videncenter for Svineproduktion, 2010. Årsbereting 2009. Avl og Genetik.Videncenter for Svineproduktion. 45 pp.3.Smith, C., Dickerson, G.E., Tess, W. and Benett, G.L., 1983. Expectedrelative responses to selection for alternative measures of life cycle economicefficiency of pork production. J. Anim. Sci. 56: 1306-1314.4.Blasco, A., Bidanel, J.P., Haley, C. 1995. Genetics and neonatal survival, pp.17-38, The Neonatal Pig. Development and Survival. Edt by M.A. Varley. CABInternational, UK.5.Bichard, M., Bovey, M., Seidel, L., David, P., Tomkins, C., 1983. NewDevelopments in Scientific Pig Breeding., No. 3, PIC, UK.6.Tomes, G.J., Nielsen, H.E. 1984. Some aspects of selection for increasedreproductive performance in sow herds, Proc. Of the 8th Int. Pig Vet. Soc.Congress, Ghent, Belgium: 203.7.Legault, C. 1985. Selection of breed strains and individual pigs for prolificacy.J. Reprod. and Fertility 33: 151-166.8.Sorensen, D., Vernersen, A., Andersen, S. 2000. Bayesian analysis ofresponse to selection: a case study using litter size in Danish Yorkshire Pigs.Genetics 156: 283-295.9.Su, G., Lund, M.S., Sorensen, D. 2007. Selection for litter size at day five toimprove litter size at weaning and piglet survival rate. J. Anim Sci. 85:1385-1392.10. Moustsen, V.A., Poulsen, H.L. 2004. Anbefalinger vedr. dimensioner påfareboks og kassesti. Videncenter for Svineproduktion Notat 0414.
51
PATTEGRISDØDELIGHED I DK
11. Andersson M., Nielsen M.B.F., Vinther J. 2010. Løsgående drægtige søer –status 2010 og forventninger til 2013. Videncenter for Svineproduktion, notat nr.1030. 3 pp.12. Schovsbo 2010. Strukturudviklingen på det danske smågrisemarked.Svineproducenten 2http://issuu.com/schovsbo/docs/strukturudvikling13. Gustafsson B. 1983. Effects of sow housing systems in practical pigproduction.Trans. Am. Soc. Agric. Eng., 26: 1181-1185.14. Anonym. Breed choices for the 21st century, International pig Topics 20:5: 17-23.http://www.positiveaction.info/PDFs/articles/pt20.5p17.pdf15. Christiansen M.G., 2010. Interpig 2008 resultater og internationalkonkurrenceevne for dansk svineproduktion i 2008 og 2009. Videncenter forSvineproduktion Notat nr. 1005. 27 pp.16. AgroSoft 2009. Produktionsdata fra Sverige sammenlignet med Finland,Norge og Danmark. Udleveret af Dansk Svineproduktion.17. Ingris 2007. Produktionsdata fra Norge. Animalia, Norsvin. Udleveret af DanskSvineproduktion.18. Pigwin sugg 2007. Produktionsdata fra Sverige. Udleveret af DanskSvineproduktion.19. IFIP-GTTT. 2009. Average National GTTT result from 1970 to 2008 in France.http://www.itp.asso.fr/eco/resultat/pdf/retro/00gttt.pdfAccessed Sep. 1, 2009.20.Data fra Canada og USAhttp://www.pigchamp.com/summary_archives.html
21. Pedersen, L.J., Berg, P, Jørgensen, G., Andersen, I.L. 2010. Neonatal piglettraits of improtance for survival in crates and indoor pens in two genetic groupsdiffering in genetic survival rate. Submiotted to J. Anim. Sci.22. Svensmark, B., Johansen, M., Bækbo, P. 2010. Dødsårsager hos pattegrise.Internt notat VSP/L&F. 3 pp.23. English, P.R., Morrison, V. 1984. Causes and prevention of piglet mortality.Pig news and information, 5(4): 369-376.24. Svendsen, J., Bengtson, A-Ch., and Svendsen, L. 1986. Occurence andcauses of traumatic injuries in neonatal pigs. Pig News and Information, 7: 159-170.25. Canario, L., Cantoni, E., Le Bihan, E., Caritez, J.C., Billon, Y., Bidanel, J.P.,Foulley, J.L. 2006. Between-breed variability of stillbirth and its relationship withsow and piglet characteristics. J. Anim Sci. 84: 3185-3196.
52
PATTEGRISDØDELIGHED I DK
26. Roehe, R., Kalm, E. 2000. Estimation of genetic and environmental riskfactors associated with pre-weaning mortality in piglets using generalized linearmixed models. Animal Science 70: 227-240.27. Akdag, F., Arslan, S., Demir. H. 2009. The Effect of Parity and Litter Size onBirth Weight and the Effect of Birth Weight Variations on Weaning Weight and Pre-Weaning Survival in Piglet. Journal of Animal and Veterinary Advances 8: 2133-213828. Johnson, R. K., Nielsen, M.K., Casey, D.S. 1999. Responses in ovulation rate,embryonal survival, and litter traits in swine to 14 generations of selection toincrease litter size. J. Anim Sci. 77: 541-557.29. Lund, M. S., Puonti, M., Rydhmer, L., Jensen, J. 2002. Relationship betweenlitter size and perinatal and pre-weaning survival in pigs. Animal Science 74: 217-222.30. Svendsen, J. 1992. Perinatal mortality in pigs. Animal Reproduction Science28: 59-67.31. Pedersen, L.J., Jørgensen, E., Heiskanen, T., Damm, B.I. 2006. Early pigletmortality in loose-housed sows related to sow and piglet behaviour and to theprogress of parturition. Appl. Anim. Behav. Sci., 96: 215-232.32. EFSA, 2007. Scientific Report on animal health and welfare aspects ofdifferent housing and husbandry systems for adult breeding boars, pregnant,farrowing sows and unweaned piglets. The EFSA Journal (2007) 572: 1-107. �European Food Safety Authority, 2007.33. Roehe, R., Shrestha, N.P., Mekkawy, W., Baxter, E.M., Knap, P.W.,Smurthwaite, K.M., Jarvis, S., Lawrence, A.B., Edwards. S.A. 2009. Geneticanalyses of piglet survival and individual birth weight on first generation data of aselection experiment for piglet survival under outdoor conditions. Livestock Science121: 173-181.34. Foxcroft, G.R., Dixon, W.T., Novak, C.T., Putman, C.T., Town, S.C., Vinsky,M.D.A. 2006. The biological basis for prenatal programming of postnatalperformance in pigs. J. Anim. Sci. 84(E.Suppl.):E105-E112.35. Wu, G., Bazer, F.W., Wallace, J.M., Spencer, T.E. 2006. Board-invited review:Intrauterine growth retardation: Implications for the animal science. J. Anim.Science. 84: 2316-2337.36. Tuchscherer, M., Puppe, B., Tuchscherer, A., Tiemann, U. 2000a. Earlyidentification of neonates at risk: Traits of newborn piglets with respect to survival.Theriogenology 54: 371-388.
53
PATTEGRISDØDELIGHED I DK
37. Milligan, B. N., Dewey, C.E., de Grau, A. F. 2002. Neonatal-piglet weightvariation and its relation to pre-weaning mortality and weight gain on commercialfarms. Preventive Veterinary Medicine 56: 119-127.38. Quiniou, N., Dagorn, J.,D. Gaudré, D. 2002. Variation of piglets' birth weightand consequences on subsequent performance. Livestock Production Science 78:63-70.39. Boulot, S., Quesnel, H., Quiniou, N. 2008. Management of high prolificacy inFrench herds: Can we alleviate side effects of piglet survival? Advances in PorkProduction. 19:1-18.40. Freking, B.A., Leymaster, K.S., Vallet, J.L., Christenson, R.K. 2007. Numberof foetuses and conceptus growth throughout gestation in lines of pigs selected forovulation rate or uterine capacity. J. Anim. Sci. 85:2093-2103.41. Edwards, S. A., Smith, W.J., Fordyce, C., Macmenemy, F. 1994. An Analysisof the Causes of Piglet Mortality in A Breeding Herd Kept Outdoors. VeterinaryRecord 135: 324-327.42. English, P.R.,Edwards, S.A. 1996. Management of the bursing sow and herlitter. In Dunkin, A.C., Taverner, M (eds) Pig Production. World Animal Science, vol.C10. Elsevier, Amsterdam: 113-140.43. Thorup, F. 1995. Besætninger med få og mange dødfødte grise. Erfaring nr.9506, Landsudvalget for Svin.44. Vaillancourt, J. P., Stein, T.E., Marsh, W.E., Leman, A.D., Dial, G.D. 1990.Validation of Producer-Recorded Causes of Preweaning Mortality in Swine.Preventive Veterinary Medicine 10: 119-130.45. Christensen, J., Svensmark, B. 1997. Evaluation of producer-recorded causesof preweaning mortality in Danish sow herds. Preventive Veterinary Medicine 32:155-164.46. Borges, V. F., Bernardi, M.L., Bortolozzo, F.P., Wentz, I. 2008. Stillbirthpattern according to birth order, birth weight, and gender of piglets. ArquivoBrasileiro de Medicina Veterinaria e Zootecnia 60: 1234-1240.47. van Dijk, A. J., van Rens, B.T.T.M., van der Lende, T., Taverne, M.A.M. 2005.Factors affecting duration of the expulsive stage of parturition and piglet birthintervals in sows with uncomplicated, spontaneous farrowings. Theriogenology 64:1573-1590.
54
PATTEGRISDØDELIGHED I DK
48. Zaleski, H. M., Hacker, R.R. 1993. Variables Related to the Progress ofParturition and Probability of Stillbirth in Swine. Canadian Veterinary Journal-RevueVeterinaire Canadienne 34: 109-113.49. Oliviero, C., Heinonen, M., Valros, A., Peltoniemi, O. 2010. Environmental andsow-related factors affecting the duration of farrowing. Animal ReproductionScience 119: 85-91.50. Baxter, E. M., Jarvis, S., Sherwood, L., Robson, S.K., Ormandy, E., Farish,M., Smurthwaite, K.M., Roehe, R., Lawrence, A.B., Edwards, S.A. 2009. Indicatorsof piglet survival in an outdoor farrowing system. Livestock Science 124: 266-276.51. Leenhouwers, J. I., Wissink, P., van der Lende, T., Paridaans, H., Knol, E.F.2003. Stillbirth in the pig in relation to genetic merit for farrowing survival. J. AnimSci. 81: 2419-2424.52. Grandinson, K., Lund, M.S., Rydhmer, L., Strandberg, E. 2002. Geneticparameters for the piglet mortality traits crushing, stillbirth and total mortality, andtheir relation to birth weight. Acta Agriculturae Scandinavica Section A-AnimalScience 52: 167-173.53. Herpin, P., Hulin, J.C., Le Dividich, J.,Fillaut, M. 2001. Effect of oxygeninhalation at birth on the reduction of early postnatal mortality in pigs. J. Anim Sci.79: 5-10.54. Leenhouwers, J. I., van der Lende, T.,Knol, E.F. 1999. Analysis of stillbirth indifferent lines of pig. Livestock Production Science 57: 243-253.55. Lucia, J., Corrêa, M.N., Deschamps, J.C., Bianchi, I., Donin, M.A., Machado,A.C., Meincke, W., Matheus, J. E. M. 2002. Risk factors for stillbirths in two swinefarms in the south of Brazil. Preventive Veterinary Medicine 53: 285-292.56. Baxter, E. M., Jarvis, S., D'Eath, R.D., , D.W., Robson, S.K., Farish, M.,Nevison, I.M., B. Lawrence, A.B., . Edwards, S.A. 2008. Investigating thebehavioural and physiological indicators of neonatal survival in pigs.Theriogenology 69: 773-783.57. Herpin, P., Damon, M., Le Dividich, J. 2002. Development of thermoregulationand neonatal survival in pigs. Livestock Production Science 78: 25-45.58. Weary, D. M., Pajor, E.A., Fraser, D., Honkanen, A.M. 1996. Sow bodymovements that crush piglets: a comparison between two types of farrowingaccommodation. Applied Animal Behaviour Science 49: 149-158.
55
PATTEGRISDØDELIGHED I DK
59. De Passille, A. M. B.,Rushen, J. 1989. Using Early Suckling Behavior andWeight-Gain to Identify Piglets at Risk. Canadian Journal of Animal Science 69:535-544.60. Blecha, F., Kelley, K.W.1981. Effects of Cold and Weaning Stressors on theAntibody-Mediated Immune-Response of Pigs. J. Anim Sci. 53: 439-447.61. Malmkvist, J., Pedersen, L.J., Damgaard, B.M.,Thodberg, K., Jorgensen, E.,Labouriau, R. 2006. Does floor heating around parturition affect the vitality of pigletsborn to loose housed sows? Applied Animal Behaviour Science 99: 88-105.62. Herpin, P., Le Dividich, J., Hulin, J.C., Fillaut, M., DeMarco, F.,Bertin, R. 1996.Effect of the level of asphyxia during delivery on viability at birth and early postnatalvitality of newborn pigs. J. Anim Sci. 74: 2067-2075.63. Holmgren, N., Lindberg, M., Linder, S., Lundeheim, N., Segall, T. 2004. Pigletmortality caused by Clostridium perfringens type A. Proceedings 18th IPVSCongress, June 27th – July 1st 2004, Hamburg, Germany 1: 260.64. Weber, R., Keil, N.M., Fehr, M., Horat, R. 2009. Factors affecting pigletmortality in loose farrowing systems on commercial farms. Livestock Science 124:216–222.65. Gardner, I.A., Hird, D.W., Franti, C.E. 1989. Neonatal Survival in Swine -Effects of Low Birth-Weight and Clinical-Disease. American Journal of VeterinaryResearch 50: 792-797.66. Sommerville, B.A., Broom, D.M. 1998. Olfactory awareness. Appl. Anim.Behav. Sci. 57: 269-286.67. Gregory, N.G. 2004. Physiology and behaviour of animal suffering. UFEW,Blackwell Science, Herts, UK, 268 s.68. Mellor, D.J., Stafford, K.J. 2004. Animal welfare implications of neonatalmortality and morbidity in farm animals. The Vet. J. 168: 118-133.69. Herskin, M.S., Jensen, K,H. 2008. Smerte og lindring heraf under og efterkastraktion af pattegrise. DJF Intern Rapport Husdyrbrug Nr. 9. Juni 2008. www.Agrsci.dk.70. Olsen, A.K., Svendsen, O. 2006. Kan fostre opleve smerte ? Danskveterinærtidsskrift nr. 15/16: 8-9.71. Busija, D.W., Leffler, C.W. 1987. Hypothermia reduces cerebral metabolic rateand cerebral blood flow in newborn pigs. Am. J. Physiol. 253: H869-H873.
56
PATTEGRISDØDELIGHED I DK
72. Gregory, N.G. 1998. Physiological mechanisms causing sickness behaviorand suffering in diseased animals. Anim. Welfare 7: 293-305.73. Su, G., Sorensen, D., Lund M.S. 2008. Variance and covariance componentsfor liability of piglet survival during different periods. Animal 2: 184-189.74. Gourdine J.L., de Greef K.H. and Rydhmer L. 2010. Breeding for welfare inoutdoor pig production: A simulation study. Livestock Science 132: 26-3475. Olesen, I., Groen A.F., Gjerde, B. 2000. Definition of animal breeding goals forsustainable production systems. J. Animal Science, 78: 570-58276. Canario L., Lundgren H., Haandlykken, M., Rydhmer L. 2010. Genetics ofgrowth in piglets and the association with homogeneity of body weight within litters.J. Anim. Sci. 88:1240-124777. Wolf, J., Zakova, E., Groeneveld, E. 2008. Within-litter variation of birth weightin hyperprolific Czech Large White sows and its relation to litter size traits, stillbornpiglets and losses until weaning. Livestock Science 115: 195-205.78. Knol, E.F., Leenhouwers, J.I., van der Lende, T. 2002. Genetic aspects ofpigletsurvival. Livestock Production Science 78: 47-5579. Nissen, P.M., Oksbjerg, N. 2009. Quantification of prenatal effects onproductivity in pigs, I Greenwood, P & Bell, A (red.), Managing prenataldevelopment to enchance livestock productivity, International Atomic EnergyAgency. IAEA, Publications, Sales and Promotion Unit: 37-69.80. Thorup, F., 2009. Effekt af B12-vitamin til drægtige søer. DanskSvineproduktion og Den rullende Afprøvning. Meddelelse nr. 850. 27 pp.81. Viberg, D. 2001. Vitaminer med henblik på svineernæring. 15-points opgave ihusdyrernæring, Den Kgl. Veterinær- og Landbohøjskole, Frederiksberg. 129pp.82. Otel, V., Costin, G, Oeriu, I. 1972. The use of folcystin for the control ofembryo mortality in pigs. II. Results of large scale experiments. Zentralbl.Veterinärmed. Reihe A 19:9: 766.83. Matte, J.J., Girard, C.L. and Brisson, G.J. 1984. Folic acid and reproductiveperformance of sows. J. Anim.Sci. 59: 1020.84. Morgan, B.L.G., Winick, M. 1978. The effect of folic acid supplementationduring pregnancy in the rat. Br. J. Nutr. 40: 529.85. Matte, J.J., Girard, C.L. 1996. Le Besoin en acide folique chez gravide,utilisation metabolique des folates comme citere de measure. Journees de laRecherche Porcine en France, nr 28: 365-370.57
PATTEGRISDØDELIGHED I DK
86. Mateo, R.D., Wu, G., Bazer, F.W., Park, J.C., Shinzato, I. 2007. Dietary L-arginin supplementation enhances the reproductive performance of gilts. J. Nutr.137: 652-656.87. Xiao, X.M., Li, L.P. 2004. L-arginine treatment for asymmetric fetal growthrestriction. Int. J. Gynecol Obstet. 2004;88:15-18.88. Mateo, R.D., Wu, G., Moon, H.K., Carroll, J.A., Kim, S.W. 2007. Effects ofdietary arginine supplementation during gestation and lactation on the performanceof lactating primiparous sows and nursing piglets. J. Anim.Sci. 86:827-835.89. Theil, P. K. 2010. Energien i råmælk er vigtig for pattegrisenes overlevelse.VilomixMagasinet, juni 2010: 4-5.90. Theil, P.K., Nielsen, M.O., Lauridsen, C., Sørensen, M.T. 2010c. Lærebog isvinets ernæring, Kapitel 16, Laktation. In Preparation.91. Jean, K.B., Chiang, S.H. 1999. Increased survival of neonatal pigs bysupplementing medium-chain triglycerides in late-gestating sow diets. Anim FeedSci Technol 76, 241-250.92. Theil, P. K., Cordero, G., Henckel, P., Puggaard, L., Oksbjerg, N., Sørensen,M.T. 2010b. Effects of gestation and transition diets, piglet birth weight and fastingtime on depletion of glycogen pools in liver and three muscles of newborn piglets.Submitted to Journal of Animal Science, januar 2010.93. Meydani, S.N., Dupont, J. 1977. Effect of zinc deficiency on prostaglandinsynthesis in different organs of the rat. J. Nutr. 112: 1098-1104.94. Kruse, P.E., Danielsen, V., Nielsen, H.E., Christensen, K. 1977. The influenceof different dietary levels of linoleic acid on reproductive performance and fatty acidcomposition of milk fat and plasma lipids in pigs. Acta Agric. Scand. 27:4: 289-296.95. Lauridsen, C., Stagsted, J., Jensen, S.K. 2007. N-6 and n-3 fatty acids ratioand vitamin E in porcine maternal diet influence the antioxidant status and immunecell eicosanoid response in the progeny. Prostaglandins and other Lipid Mediators,84: 66-78.96. Horrobin, D.F., Cunnane, S.C. 1989. Interaction between zinc, essential fattyacids and prostaglandins: relevance to acrodermatitis and entero pathica, totalparental nutrition, the glucagonoma syndrome, diabetes, anorexia nervosa andsickle cell anemia. Med. Hypothesis 6: 277-296.97. Palludan, B., Wegger, I. 1976. Studies of importance of zinc for fetaldevelopment in swine. Teratology, 13, 2: A32.
58
PATTEGRISDØDELIGHED I DK
98. Kalinowski, J., Chavez, E.R. 1984. Effect of low dietary zinc during lategestation and early lactation on the sow and neonatal piglets. Can. J. Agric. Sci. 64:749-758.99. Kalinowski, J., Chavez, E.R. 1986a. Low dietary intake during pregnancy andlactation of gilts. II. Effects on the dam. Can. J. Agric. Sci. 66: 201-216.100. Kalinowski, J., Chavez, E.R. 1986b. Low dietary intake during pregnancy andlactation of gilts. II. Effects on the offspring. Can. J. Agric. Sci. 66: 217-227.101. Lauridsen, C., Jensen, S.K. 2007. Lipid composition of lactational dietsinfluences the fatty acid profile of the progeny before and after suckling. Animal,1:7: 952-962.102. Lauridsen, C., Jensen, S.K. 2005. Influence of supplementation of all-rac-α-tocopheryl acetate preweaning and vitamin C postweaning onα-tocopherolandimmune responses of piglets. J. Anim. Sci., 83: 1274-1286.103. Omtvedt, I.T., Nelson, R.E., Edwards, R.L., Stephens, D.F., Turman, E.J.1971. Influence of heat stress during early, mid and late pregnancy of gilts. Journalof Animal Science 32(2): 312-317.104. Bate, L.A., Ireland W., Connell B.J., Grimmelt, B. 1991. Development of thesmall intestine of piglets in response to prenatal elevation of glucocorticoids.Histology and Histopathology 6:207-216.105. Haussmann, M. F., Carroll, J. A., Weesner, G. D., Daniels, M. J., Matteri R. L.,Lay, D. C. 2000. Administration of ACTH to restrained, pregnant sows alters theirpigs' hypothalamic-pituitary-adrenal (HPA) axis. J Anim Sci 78:2399-2411.106. Otten, W., Kanitz, E., Tuchscherer, M., Nürnberg, G. 2001. Effects of prenatalrestraint stress on hypothalamic-pituitaryadrenocortical and sympatho-adrenomedullary axis in neonatal pigs. Animal Science 73: 279-287.107. Kranendonk, G., Hopster, H., Fillerup, M., Ekkel, E.D., Mulder, E.J.H.,Wiegant, V.M.,Taverne, M.A.M. 2006. Lower birth weight and attenuatedadrenocortical response to ACTH in offspring from sows that orally received cortisolduring gestation. Domestic Animal Endocrinology 30: 218–238.108. Machado-Neto, R., Graves, C.N., Curtis, S.E. 1987.Immunoglobulins in pigletsfrom sows heat-stressed prepartum. J. Anim. Sci. 65:445-455.109. Tuchscherer, M., Kanitz, E., Otten, W., Tuchscherer, A. 2002. Effects ofprenatal stress on cellular and humural immune responses in neonatal pigs.Veterinary Immunology and Immunopathology, 86: 195-203.
59
PATTEGRISDØDELIGHED I DK
110. Jarvis, S., Moinard, C., Robson, S.K., Baxter, E., Ormandy, E., Douglas, A.J.,Seckl, J.R., Russell, J.A., Lawrence, A.B. 2006. Programming the offspring of thepig by prenatal social stress: Neuroendocrine activity and behaviour. Hormones andBehavior 49: 68 – 80.111. Lay, D. C., Kattesh, H.G., Cunnick, J.E., Daniels, M. J., McMunn, K. A.,Toscano, M. J., Roberts, M.P. 2008. Prenatal stress effects on pig development andresponse to weaning. J Anim Sci 86:1316-1324.112. Couret D., Otten, W., Puppe, B., Prunier, A., Merlot, E. 2009. Behavioural,endocrine and immune responses to repeated social stress in pregnant gilts. Animal3(1):118–127.113. Kanitz, E., Otten, W., Tuchscherer, M. 2006. Changes in endocrine andneurochemical profiles in neonatal pigs prenatally exposed to increased maternalcortisol. Journal of Endocrinology 191:207-220.114. Castren, H., Algers, B., Depassille, A.M., Rushen, J., Uvnasmoberg, K. 1993.Early Milk Ejection, Prolonged Parturition and Periparturient Oxytocin Release inthe Pig. Animal Production 57: 465-471.115. Oliviero, C., Heinonen, A., Valros, A., Halli, O., Peltoniemi, O.A.T. 2008. Effectof the environment on the physiology of the sow during late pregnancy, farrowingand early lactation. Animal Reproduction Science 105: 365-377.116. Lawrence, A. B., Mclean, K.A., Jarvis, S., Gilbert, C.L., Petherick, J.C. 1997.Stress and parturition in the pig. Reproduction in Domestic Animals 32: 231-236.117. Fraser, D., Phillips, P.A., Thompson, B.K. 1997. Farrowing behaviour andstillbirth in two environments: an evaluation of the restraint-stillbirth hypothesis.Applied Animal Behaviour Science 55: 51-66.118. Lawrence, A. B., Petherick, J.C., Mclean, K.A., Deans, L.A., Chirnside, J.,Vaughan, A., Clutton, E., Terlouw, E.M.C. 1994. The Effect of Environment onBehavior, Plasma-Cortisol and Prolactin in Parturient Sows. Applied AnimalBehaviour Science 39: 313-330.119. Jarvis, S., Lawrence, A.B., Mclean, K.A., Deans, L.A., Chirnside, J., Calvert,S.K. 1997. The effect of environment on behavioural activity, ACTH, beta-endorphinand cortisol in pre-farrowing gilts. Animal Science 65: 465-472.120. Damm, B. I., Lisborg, L., Vestergaard, K.S., Vanicek, J. 2003. Nest-building,behavioural disturbances and heart rate in farrowing sows kept in crates andSchmid pens. Livestock Production Science 80: 175-187.
60
PATTEGRISDØDELIGHED I DK
121. Cronin, G. M., Simpson, G.J., Hemsworth, P.H. 1996. The effects of thegestation and farrowing environments on sow and piglet behaviour and pigletsurvival and growth in early lactation. Applied Animal Behaviour Science 46: 175-192.122. Pedersen, L. J., Jensen, T. 2008. Effects of late introduction of sows to twofarrowing environments on the progress of farrowing and maternal behavior. J.Anim Sci. 86: 2730-2737.123. Biensen, N. J., von Borell, E.H., Ford, S.P. 1996. Effects of space allocationand temperature on periparturient maternal behaviors, steroid concentrations, andpiglet growth rates. J. Anim Sci. 74: 2641-2648.124. Damm, B. I., Bildsoe, M., Gilbert, C., Ladewig, J., Vestergaard, K.S. 2002a.The effects of confinement on periparturient behaviour and circulating prolactin,prostaglandin F-2 alpha and oxytocin in gilts with access to a variety of nestmaterials. Applied Animal Behaviour Science 76:135-156.125. Kristensen, A.R, Søllested, T.A. 2004. A sow replacement model usingBayesian updating in a three-level hierarchic Markov process: II. Optimizationmodel. Livestock Production Science, 87-1: 25-36.126. Da Silva A., Anil S.S., Deen J., Morrison, R.B. 2010. Claw lesion effects onpiglets born alive of stall-housed sows in two sow herds. Abstract O.261.Proceedings of the 21st IPVS Congress, Vancouver, Canada – July 18-21, 2010:293.127. Højbjerg, P.E., 2003. Urinblærebetændelse, farefeber og dødfødte grise. VestVet nyhedsbrev, 2 (3): 2.http://www.vest-vet.dk/files/nyhedsbreve/vestvet6b.pdf128. Vanderhaeghe, C., Dewulf, J., Ribbens, S., de Kruif, A., Maes, D. 2010. Across-sectional study to collect risk factors associated with stillbirths in pig herds.Animal Reproduction Science 118: 62-68.129. Christison, G. I., Wenger, I.I., Follensbee, M.E. 1997. Teat seeking success ofnewborn piglets after drying or warming. Canadian Journal of Animal Science 77:317-319.130. White, K. R., Anderson, D.M., and Bate, L.A. 1996. Increasing piglet survivalthrough an improved farrowing management protocol. Canadian Journal of AnimalScience 76: 491-495.131. Andersen, I. L., Haukvik, I.A., Boe, K.E. 2009. Drying and warmingimmediately after birth may reduce piglet mortality in loose-housed sows. Animal 3:592-597
61
PATTEGRISDØDELIGHED I DK
132. Thorup, F., Herskin, M.S., Hansen, E.N., Musse, S.L., Nielsen, J.P., Schmidt,M.H., 2009. Faringsforløb hos frugtbare danske søer. Medd 839.http://vsp.lf.dk/Publikationer/Kilder/lu_medd/2009/839.aspx133. Thorup, F., 2005. Faringsinduktion. Dansk Svineproduktion,http://vsp.lf.dk/Viden/Reproduktion/Faring_diegivning/Faringsinduktion.aspx134. Papadopoulos, G. A., Vanderhaeghe,C., Janssens, G.P.J., Dewulf, J., Maes,D.G.D. 2010. Risk factors associated with postpartum dysgalactia syndrome insows. Veterinary Journal 184: 167-171.135. Farmer, C., Quesnel, H. 2009. Nutritional, hormonal, and environmentaleffects on colostrum in sows. J Anim Sci. 87:56-64.136. Pedersen, B.K. 1986. Afprøvning af faringsinduktion. Landsudvalget for svinog Den rullende Afprøvning. Meddelelse nr. 112, 7 pp.137. Algers, B., Jensen, P. 1990. Thermal Microclimate in Winter Farrowing Nestsof Free-Ranging Domestic Pigs. Livestock Production Science 25: 177-181.138. Hrupka, B. J., Leibbrandt, V.D., Crenshaw, T.P., Benevenga, N.J. 1998. Theeffect of farrowing crate heat lamp location on sow and pig patterns of lying and pigsurvival. J. Anim Sci. 76: 2995-3002.139. Vasdal, G., Andersen, I.L., Pedersen, L.J. 2009. Piglet use of the creep area -Effects of breeding value and farrowing environment. Applied Animal BehaviourScience 120: 62-67.140. Vasdal, G., Glaerum, M., Melisova, M., Boe, K.E., Broom, D.M., I. L.Andersen, I.L. 2010. Increasing the piglets' use of the creep area-A battle againstbiology? Applied Animal Behaviour Science 125: 96-102.141. Gu, Z. B., Xin, H.W., Wang, C.Y.,Shi, Z.X., Liu, Z.H., Yang, F.Y.,Lin, B.Z.,Wang, C., Li, B.M. 2010. Effects of neoprene mat on diarrhea, mortality and forelegabrasion of pre-weaning piglets. Preventive Veterinary Medicine 95: 16-22.142. Pedersen, B.K. 1988. Varmelamper i farestalde, Infosvin, Medd. 138.143. Algers, B., Jensen, P. 1991. Teat Stimulation and Milk-Production DuringEarly Lactation in Sows - Effects of Continuous Noise. Canadian Journal of AnimalScience 71:51-60.144. Nielsen, B.L., Thodberg, K., Dybkjær, L., Vestergaard, E.M. 2006. Feedingbehaviour in Pigs. In “Feeding in domestic vertebrates. From structure to behaviour”(eds V. Bels). CABI publishing, Wallingford, UK: 156-178.
62
PATTEGRISDØDELIGHED I DK
145. Algers, B. 1993. Nursing in Pigs - Communicating Needs and DistributingResources. J. Anim Sci. 71: 2826-2831.146. Fraser, D., Thompson, B.K.1986. Variation in Piglet Weights - Relationship toSuckling Behavior, Parity Number and Farrowing Crate Design. Canadian Journalof Animal Science 66: 31-46.147. Moustsen, V.A., Poulsen, H.L. 2004a.’Pattegrises dimensioner’, Notat 0432,Landsudvalget for Svin.148. Moustsen, V.A. 2006. Sammenligning af to dimensioner af kassestier. Medd.739, Dansk svineproduktion.149. Moustsen, V.A., Poulsen, H.L. 2004b. Sammenligning af produktionsresultateropnået i henholdsvis en traditionel kassesti og en sti til løsgående farende ogdiegivende søer. Meddelelse 679. Landsudvalget for svin.150. Algers, B., Jensen, P. 1985. Communication During Suckling in the DomesticPig - Effects of Continuous Noise. Applied Animal Behaviour Science 14: 49-61.151. De Passille, A. M. B., Rushen, J., Hartsock, T.G. 1988. Ontogeny of TeatFidelity in Pigs and Its Relation to Competition at Suckling. Canadian Journal ofAnimal Science 68: 325-338.152. Le Dividich, J., Rooke, J.A., Herpin, P. 2005. Nutritional and immunologicalimportance of colostrum for the new-born pig. Journal of Agricultural Science 143:469–485.153. Thorup, F. 2010. 11,13 eller 15 diende grise hos soen. Medd 872.http://vsp.lf.dk/Publikationer/Kilder/lu_medd/2010/872.aspx154. Neal, S.M., Irvin, K.M. 1991. The effects of crossfostering pigs on survival andgrowth. J. Anim. Sci. 69: 41-46.155. Price, E. O., Hutson, G.D., Price, M.I., Borgwardt, R. 1994. Fostering in SwineAs Affected by Age of Offspring. J. Anim Sci. 72: 1697-1701.156. Pedersen, L.J., Jensen, H., Thodberg, K. 2008. Cross fostering of piglets infarrowing pens. Proceedings of the 20th Nordic Symposium of the InternationalSociety for Applied Ethology.157. Horrell, I., Bennett, J. 1981. Disruption of Teat Preferences and Retardation ofGrowth Following Cross-Fostering of 1-Week-Old Pigs. Animal Production 33: 99-106.158. Horrell, R. I. 1982. Immediate Behavioral Consequences of Fostering 1-Week-Old Piglets. Journal of Agricultural Science 99: 329-336.63
PATTEGRISDØDELIGHED I DK
159. Wattanakul, W., Stewart, A.H., Edwards, S.A., English, P.R. 1998. The effectof cross-suckling and presence of additional piglets on sucking behaviour andperformance of individually housed litters. Animal Science 66: 449-455.160. Stewart, T. S., Diekman, M.A. 1989. Effect of Birth and Fraternal Litter Sizeand Cross-Fostering on Growth and Reproduction in Swine. J. Anim Sci. 67: 635-640.161. Straw, B. E., Burgi, E.J., Dewey, C.E., Duran, C.O. 1998a. Effects ofextensive crossfostering on performance of pips on a farm. Journal of the AmericanVeterinary Medical Association 212: 855-+.162. Giroux, S., Robert, S., Martineau, G.P. 2000. The effects of cross-fostering ongrowth rate and post-weaning behavior of segregated early-weaned piglets.Canadian Journal of Animal Science 80: 533-538.163. Straw, B. E., Dewey, C.E., Burgi, E.J.1998b. Patterns of crossfostering andpiglet mortality on commercial US and Canadian swine farms. Preventive VeterinaryMedicine 33: 83-89.164. Thorup, F. 1998. Kuldudjævningens betydning for fravænningsvægten.Erfaring 9804.http://vsp.lf.dk/Publikationer.aspx165. Thorup, F.1999b, Afprøvning af strategier for kuldudjævning. Medd. nr.453.http://vsp.lf.dk/Publikationer/Kilder/lu_medd/2010/872.aspx166. Aherne, 1999, International Pig Letter: 4-6.167. De Passille, A. M. B., Robert, S. 1989. Behavior of Lactating Sows - Influenceof Stage of Lactation and Husbandry Practices at Weaning. Applied AnimalBehaviour Science 23: 315-329.168. Götz, M. 1991. Changes in nursing and suckling behaviour of sows and theirpiglets in farrowing crates. Appl Anim Behav Sci 31: 271-275.169. Boe, K. 1991. The Process of Weaning in Pigs - When the Sow Decides.Applied Animal Behaviour Science 30: 47-59.170. Damm, B. I., Pedersen, L.J., Jessen, L.B., Thamsborg, S.M., Mejer, H.,Ersboll, A.K. 2003. The gradual weaning process in outdoor sows and piglets inrelation to nematode infections. Applied Animal Behaviour Science 82: 101-120.171. Pitts, A. D., Weary, D.M., Fraser, D., Pajor, E.A., Kramer, D.L. 2002.Alternative housing for sows and litters Part 5. Individual differences in the maternalbehaviour of sows. Applied Animal Behaviour Science 76: 291-306.
64
PATTEGRISDØDELIGHED I DK
172. Cronin, G. M., Barnett, J.L.,Hodge, F.M., Smith, J.A., McCallum, T.H. 1991.The welfare of pigs in two farrowing/lactation environments: cortisol responses ofsows. Applied Animal Behaviour Science 32: 117-127.173. Jarvis, S., D'Eath, R.B., Robson, S.K., Lawrence, A.B. 2006. The effect ofconfinement during lactation on the hypothalamic-pituitary-adrenal axis andbehaviour of primiparous sows. Physiology & Behavior 87: 345-352.174. Weary, D. M., Pajor, E.A., Bonenfant, M., Fraser, D., Kramer, D.L. 2002.Alternative housing for sows and litters.: Part 4. Effects of sow-controlled housingcombined with a communal piglet area on pre- and post-weaning behaviour andperformance. Applied Animal Behaviour Science 76: 279-290.175. Thorup, F., Callesen, J., Udesen, F.K., 2006. Økonomisk betydning af 4 eller 5ugers fravænning. Meddelelse nr. 759. Dansk Svineproduktion, Den rullendeAfprøvning.http://vsp.lf.dk/Publikationer.aspx176. Pajor, E. A., Weary, D.M.,Caceres, C., Fraser, D., and Kramer, D.L. 2002.Alternative housing for sows and litters Part 3. Effects of piglet diet quality and sow-controlled housing on performance and behaviour. Applied Animal BehaviourScience 76: 267-277.177. Bonde, M.K., Herskin, M.S., Jørgensen, E., Jensen, K.H. 2007. Vidensynteseom skuldersår. DJF rapport. Pp. 105.178. Long, T., Aasmundstad, T., Holm, B. 2010. Genetics of teat quantity andquality in Norwegian Landrace. 9th World Congress on Genetics Applied toLivestock Production.179. Kelly, D., Coutts, A.G.P. 2000. Development of digestive and immunologicalfunction in neonates: role of early nutrition. Livestock Production Science 66: 161–167.180. Rooke, J.A., Bland I.M. 2002. The acquisition of passive immunity in the new-born piglet. Livestock Production Science 78: 13–23.181. Jourquin, J., Biermann, J., van Gelderen, R., Groossens, L. 2010. Bettercocostrum distribution increases piglet survival in high prolific sows. Abstract O.262.Proceedings of the 21st IPVS Congress, Vancouver, Canada – July 18-21, 2010, p.294.182. Thorup, F. Ebbesen, T. 2009. Farefeber / MMA / mastitis, metritis og agalaksi.Dansk Svineproduktion,http://www.infosvin.dk/Haandbog/Sundhed/Sygdomme_lidelser/Koensdele_koensveje/Farefeber%20MMA%20mastitis%20metritis%20og%20agalaksi.html
65
PATTEGRISDØDELIGHED I DK
183. Devillers, N., Farmer C., Le Dividich J., Prunier, A. 2007. Variability ofcolostrum yield and colostrum intake in pigs. Animal 1(7): 1033–1041.184. Foisnet, A., Farmer, C., David C., Quesnel, H. 2010. Relationships betweencolostrum production by primiparous sows and sow physiology around parturition. JAnim Sci 88: 1672-1683.185. Azain, M.J. (1993) Effects of adding medium-chain triglycerides to sow dietsduring late gestation and early lactation on litter performance. J Anim Sci 71: 3011-3019.186. Nissen, S., Faidley, T.D., Zimmerman, D..R, Izard, R.,Fisher, C.T. 1994.Colostral milk fat percentage and pig performance are enhanced by feeding theleucine metabolite beta-hydroxy-beta-methyl butyrate to sows. J Anim Sci 72: 2331-2337.187. Theil, P. K., Hansen, A.V., Sørensen, M.T. 2010a. Effects of gestation andtransition diets on colostrum intake and piglet survival. Poster presented at the 61stEAAP meeting, 23-27 August 2010, Crete.188. Lauridsen, C., Danielsen, V. 2004. Lactational dietary fat levels and sourcesinfluence milk composition and performance of sows and their progeny. Livest.Prod. Sci. 91:95-105.189. Odle, J., Benevenga, N.J., Crenshoaw, T.D. 1992. Evaluation of [1-14C]-medium chain fatty acid oxidation by neonatal piglets using continous infutionradiotracer kinetic methodology. J. Nutr. 122: 2183-2189.190. Bækbo, P. 2006. Immunitet/immunitetsstyring.http://www.infosvin.dk/Haandbog/Sundhed/Generelt_sundhed/Immunitet_immunitetsstyring.html191. Christensen, P.T., 2008. Vacciner og –programmer.http://www.infosvin.dk/Haandbog/Sundhed/Generelt_sundhed/Vacciner.html192. Jorsal, S.E. 1983. Morbiditet hos søer. Epidemiologiske undersøgelser iintensive sobesætninger med særligt henblik på farefebersyndromet.Licentiatafhandling, Institut for intern medicin, Den kgl. Veterinær- ogLandbohøjskole, København. 118 pp.193. Salwén, T., Wallgren, P. 2010. Time point for vaccination sows with E.colivaccines may influence the protection of the offspring. Abstract O.052. Proceedingsof the 21st IPVS Congress, Vancouver, Canada – July 18-21, 2010, p. 90.194. Svensmark, B. 2009. New neonatal diarrhoea syndrome in Denmark.Proceedings of the 1st ESPHM, Copenhagen, Denmark, 2009, p. 27.
66
PATTEGRISDØDELIGHED I DK
195. Gin T., Le Guennec, J., Morvan, H., Martineau, G.P. 2010. Clinical andlaboratory investigations in 10 French pig herds dealing with enzootic neonataldiarrhea. Abstract P.452. Proceedings of the 21st IPVS Congress, Vancouver,Canada – July 18-21, 2010: 758.196. Melin, L., Wallgren, P., Mattsson, S., Stampe, M., Löfstedt, M. 2010.Neonataal diarrhoea in piglets from E.coli vaccinated sos in Sweden. AbstractO.258. Proceedings of the 21st IPVS Congress, Vancouver, Canada – July 18-21,2010, p. 290.197. Sialelli, J.-N., Vautrin, F., Lautrou, Y., Oswald, I., Quiniou, N., Martineau, G.P.2010. Farrowing progress (chronopart) and enzootic neonatal diarrhea:observational study in nine commercial herds. Abstract P.446. Proceedings of the21st IPVS Congress, Vancouver, Canada – July 18-21, 2010, p. 752.198. Bækbo P., pers. Medd., 2010.199. Krakowski, L., Krzyzanowski, J., Wrona, Z., Kosto, K., Siwicki, A.K. 2002. Theinfluence of non-specific immunostimulation of pregnant sows on the immunologicalvalue of colostrums. Veterinary Immunology and Immunophathology 87: 98-95.200. Salmon, H., Berri, M., Gerdts, V., Meurens, F. 2009. Humoral and cellularfactors of maternal immunity in swine. Developmental and ComparativeImmunology 33: 384–393.201. O’Quinn, P.R., Funderburke, D.W., Tibbetts, G.W. 2001. Effects of dietarysupplementation with mannan oligosaccharides on sow and litter performance in acommercial production system. J. Anim. Sci. 79(Suppl.1):212 (abstract).202. Alexopoupus, C., Georgoulakis I.E., Tzivara A., Kritas S.K., Siochu A.,Kyriakis, S.C. 2004. Field evaluation of the efficacy of a probiotic containing Bacilluslicheniformis and Bacillus subtilis spores, on the health status and performance ofsows and their litters. J. Anim. Physiol. a. Anim. Nutr. 88: 381–392.203. Sørensen, G. 1997. BioPlus 2B i foder til søer og pattegrise. Videncenter forSvineproduktion, meddelelse nr. 350.204. Svendsen, L.S., Weström, B.R., Svendsen, J., Olsson, A.-Ch., Karlsson, B.W.1990. Intestinal macromolecular transmission in underprivileged and unaffectednewborn pigs: implication for survival of underprivileged pigs. Research omVeterinary Science 48: 184-189.205. Xu, R.-J., Doan, Q.C., Regester, G.O. 1999. Detection and Characterisation ofTransforming Growth Factor-Beta in Porcine Colostrum. Biol Neonate 75: 59–64.
67
PATTEGRISDØDELIGHED I DK
206. Wagstrom, E.A., Yoon K.-J., Zimmerman, J.J. 2000. Immune components inporcine mammary sections. Viral Immunology 13(3): 383-397.207. Nguyen, T.V., Yuan, L., Azevedo, M.S.P., Jeong, K.-I., Gonzalez, A.-M., Saif,L.J. 2007. Transfer of maternal cytokines to suckling piglets: In vivo and in vitromodels with implications for immunomodulation of neonatal immunity. VeterinaryImmunology and Immunopathology 117: 236–248.208. Tuboly, S., Bernath, S., Glavits, R., Medveczky, I. 1988. Intestinal Absorptionof Colostral Lymphoid Cells in Newborn Piglets. Veterinary Immunology andImmunopathology, 20: 75-85.209. Williams, P.P. 1993. Immunomodulating effects of intestinal absorvedmaternal colostral leukocytes by neonatal pigs. Can. J. Vet. Res. 57: 1-8.210. Bäckström, L., Morkoc, A.C., Connor, J., Larson, R., Price, W. 1984. Clinicalstudy of mastitis-metritis-agalactia in sows in Illinois. Journal of American VeterinaryMedicine Association, 185: 70-73.211. Hansen, L.L., Vestergaard, K. 1984. Tethered versus loose sows: Ethologicalobservations and measures of productivity: II. Production results. Ann. Rech. Vét.15(2): 185-191.212. Landsudvalget for svin 1990. Transpondersystemer til drægtige søer. DanskeSlagterier, København, Meddelelse 181, 11 pp.213. Spicer, E.M., Driesen, S.J., Fahy, V.A., Horton, B.J., Sims, L.D., Jones, R.T.,Cutler, R.S., Prime, R.W. 1986. Causes of preweaning mortality on a large intensivepiggery. Australian Veterinary Journal 63(3): 71-75.214. Andersen, I.L., Berg, S., Bøe, K.E. 2005. Crushing of piglets by the mothersow (Sus scrofa) – purely accidental or a poor mother. Appl. Anim. Behav. Sci., 93:229-243.215. Thodberg, K., Jørgensen, E., Olsson, A.C., Houbak, B., Pedersen, L.J. 2008.The relationship between pre-pubertal inactivity and maternal ability in loose-housed sows. Proceedings of the 42nd Congress of the ISAE, Dublin, Ireland: 79.216. Wechsler, B., Hegglin, D. 1997. Individual differences in the behaviour of sowsat the nest-site and the crushing of piglets. Appl. Anim. Behav. Sci., 51: 39-49.217. Pokorna, Z., Illmann, G., Simeckova, M., Chaloupkova, H., Kratinova., P.2008. Carefulness and flexibility of lying down behaviour in sows during 24 h post-partum in relation to piglet position. Appl. Anim. Behav. Sci., 114: 346-358.
68
PATTEGRISDØDELIGHED I DK
218. Johnson, A. K., Morrow, J.L., Dailey, J.W., McGlone, J.J. 2007. Preweaningmortality in loose-housing lactating sows: Behavioural and performance differencesbetween sows who crush or do not crush. Appl. Anim. Behav. Sci., 105: 59-74.219. Wischner, D., Kemper, N., Stamer, E., Hellbrügge, B., Presuhn, U., Krieter, J.2010. Pre-lying behaviour patterns in confined sows and their effects on crushing ofpiglets. Appl. Anim. Behav. Sci., 122: 21-27.220. Held, S., Mason, G., Mendl, M. 2006. Maternal responsiveness of outdoorsows from first to fourth parities. Appl. Anim. Behav. Sci., 98: 216-233.221. Thodberg, K., Jensen., K.H., Herskin, M.S. 2002a. Nest building and farrowingin sows: relation to the reaction pattern during stress, farrowing environment andexperience. Appl. Anim. Behav. Sci., 77: 21-42.222. Thodberg, K., Jensen, K.H., Herskin, M.S., 2002b. Nursing behaviour,postpartum activity and reactivity in sows. Effects of farrowing environment,previous experience and temperament. Appl. Anim. Behav. Sci., 77: 53-76.223. Janczak, A.M., Pedersen, L.J., Rydhmer, L., Bakken, M. 2003. Relationbetween early fear- and anxiety-related behaviour and maternal ability in sows.Applied Animal Behaviour Science 82: 121-135.224. Grandinson, K., Rydhmer, L., Strandberg, E., Thodberg, K. 2003. Geneticanalysis of on-farm tests of maternal behaviour in sows. Livestock ProductionScience, 83: 141-151.225. Gäde, S., Bennewitz, J., Kirchner, K., Looft, H., Knap, P.W., Thaller, G., Kalm,E. 2008. Genetic parameters for maternal traits in sows. Livestock Science, 114:31-41.226. Hellbrügge, B., Tölle, K.-H., Bennewitz, J., Henze, C., Presuhn, U., Krieter, J.2008. Genetic aspects regarding piglet losses and the maternal behaviour of sows.Part 2: Genetic relationship between maternal behaviour in sows and pigletmortality.Animal, 2: 1281-1288.227. Vangen, O., Holm, B., Valros, A., Lund, M.S., Rydhmer, L. 2005. Geneticvariation in sows’ maternal behaviour, recorded under field conditions. LivestockProduction Science, 93: 63-70.228. Wischner, D., Kemper, N., Krieter, J. 2009. Nest-building behaviour in sowsand consequences for pig husbandry. Livestock Science 124:1-8.229. Burri, M., Wechsler, B., Gygax, L., Weber, R. 2009. Influence of straw length,sow behaviour and room temperature on the incidence of dangerous situations for
69
PATTEGRISDØDELIGHED I DK
piglets in a loose farrowing system. Applied Animal Behaviour Science 117: 181-189.230. Thodberg, K., Jensen, K.H., Herskin, M., Jørgensen, E. 1999. Influence ofenvironmental stimuli on nest building and farrowing behaviour in domestic sows.Appl. Anim. Behav. Sci. 63: 2: 131-144.231. Cronin, G.M., Schirmer, B.N., McCallum, T.H., Smith, J.A., Butler, K.L. 1993.The Effects of Providing Sawdust to Pre-Parturient Sows in Farrowing Crates onSow Behavior, the Duration of Parturition and the Occurrence of Intra-PartumStillborn Piglets. Applied Animal Behaviour Science 36: 301-315.232. Wischner, D., Kemper, N., Stamer, E., Hellbruegge, B., Presuhn, U., Krieter,J. 2009. Characterisation of sows' postures and posture changes with regard tocrushing piglets. Applied Animal Behaviour Science 119: 49-55.233. Edwards, S. A., Furniss, S.J. 1988. The Effects of Straw in Crated FarrowingSystems on Peripartal Behavior of Sows and Piglets. British Veterinary Journal 144:139-146.234. Herskin, M.S., Jensen, K.H., Thodberg, K. 1998. Influence of environmentalstimuli on maternal behaviour related to bonding, reactivity and crushing of pigletsin domestic sows. Appl. Anim. Behav. Sci., 58: 241-254.235. Damm, B.I., Heiskanen, T., Pedersen, L.J., Jorgensen, E., Forkman, B. 2010.Sow preferences for farrowing under a cover with and without access to straw.Applied Animal Behaviour Science 126: 97-104.236. Cronin, G.M. Vanamerongen. G. 1991. The Effects of Modifying the FarrowingEnvironment on Sow Behavior and Survival and Growth of Piglets. Applied AnimalBehaviour Science 30: 287-298.237. Pedersen, L.J., Damm, B.I., Marchant-Forde, J.N., Jensen, K.H. 2003. Effectsof feed-back from the nest on maternal responsiveness and postural changes inprimiparous sows during the first 24 h after farrowing onset. Applied AnimalBehaviour Science 83: 109-124.238. Pedersen, L.J. 2008. Important pen features and management in farrowingpens for loose housed sows. Non crate systems for non lactating sows. Internrapport Århus universitet, DJF. Seminar held in Copenhagen 12th of june 2008. pp19.239. Rasmussen, H.M.,Petersen, L.B. 2008. Redebygningsmateriale i farestier.Medd. 829.http://vsp.lf.dk/Publikationer/Kilder/lu_medd/2008/829.aspx
70
PATTEGRISDØDELIGHED I DK
240. Damm, B. I., Pedersen, L.J., Heiskanen, T., Nielsen, N.P. 2005. Long-stemmed straw as an additional nesting material in modified Schmid pens in acommercial breeding unit: effects on sow behaviour, and on piglet mortality andgrowth. Applied Animal Behaviour Science 92: 45-60.241. Jarvis, S., Reed, B.T., Lawrence, A.B., Calvert, S.K., Stevensen, J. 2004.Perinatal environmental effects on maternal behaviour, pituitary and adrenalactivation and the progress of parturition in the primiparous sows. Animal welfare13:171-181.242. Damm, B. I., Forkman, B., Pedersen, L.J. 2005. Lying down and rollingbehaviour in sows in relation to piglet crushing. Applied Animal Behaviour Science90: 3-20.243. Weber, R., Keil, N.M., Fehr, M., Horat, R. 2007. Piglet mortality on farms usingsystems with and without crates. Anim. Welfare 16: 277-279.244. Cronin, G. M., Lefébure, B., McClintock, S. 2000. A comparison of pigletproduction and survival in the Werribee Farrowing Pen and conventional farrowingcrates at a commercial farm. Australian Journal of Experimental Agriculture 40: 17-23.245. Bille, N., Nielsen, N.C., Svendsen, J. 1974. Preweaning mortality in pigs. 3.Traumatic injuries. Nord Vet.-Med. 26: 617-625.246. Nielsen, N.C., Bille, N., Svendsen, J., Riising, H.-J. 1976.Sygdomsbekæmpelse i svinebesætninger. Institut for Intern Medicin, Den kgl.Veterinær og Landbohøjskole, Frederiksberg Bogtrykkeri. 403 pp.247. Anil L., Anil S.S., Baidoo, S.K., Deen J., Wilson M.E., Ward, T.L. 2010.Factors associated with pre-weaning piglet mortality due to crushing. AbstractO.260. Proceedings of the 21st IPVS Congress, Vancouver, Canada – July 18-21,2010: 292.248. Revilla, E., Ubiergo, A., Martinez, P., Ruiz, J., Rubio, S., Salleras, J.M. 2006.Post-farrowing treatment of sows with Meloxicam on the preweaning weight gainand mortality rate of the low birth weight piglets in subclinical MMA. Abstract P.32-04. Proceedings of the 19th IPVS Congress, Copenhagen, Denmark 2006, 2: 475.249. Busch, M.E. 2009. Smertestillende behandling.http://vsp.lf.dk/Viden/Sygdom%20og%20behandling/Behandling/Smertebehandling.aspx250. Solivwww.soliv.dk
71
PATTEGRISDØDELIGHED I DK
251. Vestergaard, K., Hansen, L.U., Nielsen, E.O., Nissen, H., Sørensen, G.,Thorup, F., Kristensen, H., Studnitz, M., Bruun, L.K., Ruby, V., Christiansen, M.G.2010. Sow Life� – demonstrating the quality of advisory teams and reducing sowmortality on Danish farms. Abstract O.180. Proceedings of the 21st IPVS Congress,Vancouver, Canada – July 18-21, 2010: 214.252. Zoric, M., Nilsson, E., Mattsson, S., Lundeheim, N., Wallgren, P. 2008.Abrasions and lameness in piglets born in different farrowing systems with differenttypes of floor. Acta Veterinaria Scandinavica 50.253. Zoric, M., Nilsson, E., Lundeheim, N., Wallgren, P. 2009. Incidence oflameness and abrasions in piglets in identical farrowing pens with four differenttypes of floor. Acta Veterinaria Scandinavica 51.254. Norring, M., Valros, A., Munksgaard, L., Puumala, M., Kaustell, K.O.,Saloniemi, H. 2006. The development of skin, claw and teat lesions in sows andpiglets in farrowing crates with two concrete flooring materials. Acta AgriculturaeScandinavica Section A-Animal Science 56: 148-154.255. Lewis, E., Boyle, L.A., O'Doherty, J.V.,Brophy, P., Lynch, P.B. 2005. Theeffect of floor type in farrowing crates on piglet welfare. Irish Journal of Agriculturaland Food Research 44: 69-81.
72
PATTEGRISDØDELIGHED I DK
OrdlisteAmmeso: Diegivende so, der passer et hold fremmede grise, efter at hendes egne grise erfravænnetantioxidativ kapacitet: evnen til at modstå oxidativt stress. Oxidativt stress kan opstå vedstor aktivitet i stofskiftet, hvis den antioxidativ kapacitet er lille, og er ødelæggende forceller og væv i organismenAvlsindex: Det kriterie grisen udvælges til avl på. Dette er et vægtet gennemsnit af deestimerede avlsværdier for de egenskaber der indgår i avlsmålet, vægtet med deresrelative betydning (kr per enheds ændring). Avlsindekset beskriver dyrets værdi somforælder til næste generation.Arvbarhed: Andelen af den samlede variaton der skyldes additive genetiske effekter. Jostørre arvbarhed jo større er sammenhæng mellem fænotype og genotypeBeta-oxidation: Forbrænding af fedtstofferBioaktive komponenter: Indholdsstoffer i f.eks. foder, der har en markant og ofte unikbiologisk effekt.Drægtighedsstald: Staldafsnit i svinestald, hvor søerne er opstaldet under deresdrægtighed fra senest 4 uger efter de er insemineret og indtil få dage før forventet fødselaf grisene.Embryodannelse: Udvikling af befrugtede æg til fostre gennem celledeling ogspecialisering af cellerFarefeber: Sygdomskompleks hos so lige efter faring med feber og symptomer pålivmoderbetændelse, yverbetændelse og nedsat mælkeproduktionFarestald: Staldafsnit i svinestald, hvor søerne opstaldes fra få dage før fødsel af griseneog indtil fravænning. Søerne er typisk i farestalden i 4-5 uger, hver gang de skal fødegrise.Faringsinduktion: Igangsættelse af fødsel ved indgivelse af medikament.Fødselsinterval: Tidsrum mellem fødsel af to grise under et fødselsforløb.Generaliseret infektion: Sygdom der påvirker hele organismen (gennem feber eller flereorganer eller kropsområder er angrebet) i modsætning til en lokaliseret afgrænsetinfektion, f.eks. en byld.Genetisk korrelation: Sammenhæng mellem to egenskaber, der skyldes genetiskeeffekter.
73
PATTEGRISDØDELIGHED I DK
Genomisk selektion: Selektion i to trin, hvor man først etablerer en sammenhæng mellemgenetiske markører og fænotype og dernæst kan estimere et dyrs avlsværdi alene udfradets genetiske markører.Glykogendepot: Kulhydrat lagret i lever og muskler, der hurtigt kan spaltes til sukker ogbruges som energikilde.Gylt: Ung so der er drægtig med sit første kuld grise.Heritabilitet: se arvbarhedHomeostase: Opretholdelse af et stabilt niveau. Bruges f.eks. om regulering af blodetsindhold af glukose.Ihjellægning: Dødsfald af gris forårsaget af, at soen har lagt sig på den, og dermed klemtden. Dødsårsag enten kvælning eller på grund af traume.Immunitet: Modstandskraft mod infektioner fra f.eks. bakterier eller virusImmunkompetence: Immunsystemets evne til at reagere hensigtsmæssigt på en infektionImmunmodulerende effekt: påvirkning af immunsystemetImmunologisk kvalitet: indhold af stoffer, der fremmer modstandsdygtigheden overforsygdommeImmunstimulator: et stof, der øger immunsystemets reaktionsevneImmunsupprimerende: undertrykkelse af immunsystemetKolostrum: se RåmælkLaktat: mælkesyreLøbningstidspunkt: Tidspunkt hvor soen er i brunst og med succes kan insemineres ogblive drægtig.Mitochondrier: Cellestrukturer der sørger for forbrænding af næringsstoffer.NSAID midler: Gruppe af lægemidler der virker feber-nedsættende, smertelindrende oghæmmer betændelsestilstande.Ovulationsrate: Antal ægløsninger under en sos brunst.Patogen: Sygdomsfremkaldende mikroorganisme.Patteorden: I et kuld grise har hver gris sin faste patte, som den dier fra hver gang.Etableres i grisenes første levedøgn.
74
PATTEGRISDØDELIGHED I DK
Placenta: Vævslag i soens livmoder (moderkage) hvorigennem grisene ernæres ifostertilstanden.Polyaminer: organiske stoffer, som bl.a. menes at have betydning for regulering afcellevækstProbiotika: Ordet probiotika kommer fra græsk, hvor ”pro-” betyder til fordel for, og ”bios”oversættes til liv. Probiotika kan defineres som ”levendemikroorganismer som tilført ipassende mængder udfører en sundhedsfremmende effekt på værten”.Probiotika er somregel mikroorganismer, der findes i den naturlige mikroflora.Prostaglandin: en gruppe fede syrer, som påvirker en lang række fysiologiske processer,og som bl.a. indgår som igangsætter af fødslen.Reproduktion: Formering, frugtbarhed, produktion af afkom.Råmælk: Den særlige mælk (også kaldet kolostrum) som udskilles fra yveret under og idet første døgn efter faring. Karakteristisk for råmælk er bl.a. et højt indhold af antistoffer.Selektionsindex: Et vægtet sum af avlsværdien for forskellige egenskaber for et dyr.Vægtning foretages ud fra egenskabens marginale betydning under danske forhold udtrykti forhold til indtjening pr. produceret kilo kød. Dyrene rangeres ud fra selektionsindeksetved udvælgelse af de bedste dyr.Selektionskriterium: Normalt identisk med selektionsindeks, og benyttes oftest når kun énegenskab indgår i indeks.Termiske forhold: Temperaturmæssige forhold.Uterus: Soens livmoderÆtiologi: årsag til sygdom
75
PATTEGRISDØDELIGHED I DK
BilagBilag 1.Notat vedrørende størrelsen af farestier i 84 danske besætninger. Af MarianneBonde HBS, 29-09-2010.
Farestier i besætninger der deltog i skuldersårsprojektet 2008.I skuldersårsprojektet 2008 deltog 98 sobesætninger. Besætningerne var karakteriseretved at have min. 100 årssøer med de diegivende søer opstaldet i traditionelle kassestier,mens de drægtige søer var opstaldet i forskellige former for løsdrift-systemer.Besætningerne var lokaliseret over hele Danmark og var rekrutteret gennem henvendelsetil ca. 1000 besætninger, der var tilfældigt udvalgt fra CHR-registeret blandt besætningermed indberetning af mere end 100 voksne dyr. 247 besætninger opfyldte kravet tilopstaldning og var interesseret i at deltage i projektet. Ud fra disse blev 120 besætningertilfældigt udvalgt, og 98 besætninger heraf indgik i projektet.Der foreligger komplette data fra 84 besætninger vedr. registreringer af system ifarestalden (fra i alt 214 staldsektioner). Systembeskrivelsen inkluderer: Længde ogbredde af faresti (totalt stiareal), areal af fast gulv og spaltegulv i stien, spaltemateriale,bredde af farebøjle v. hoved, længde af farebøjle (fra bagkant krybbe), højde afstiskillevægge, strøelse, vurdering af støjniveau og støjkilder. Derudover ligger data vedr.den indstillelige bredde i bagenden af farebøjlen på en stikprøve af ca. 40 søer pr.besætning i hver af de 98 deltagende besætninger. Datamaterialet er indsamlet i periodenfebruar-august 2008.I opgørelsen nedenfor indgår hver sektion med 1 sti uafhængigt af antal stier med sammeudformning i den pågældende sektion og dermed deres ”belastning” af sopopulationen.Median og 25/75 % fraktiler angiver således ikke absolutte forhold oplevet afsopopulationen, men er ment som en angivelse af, hvilken variation i sti-dimensionering,der ses i private sobesætninger. Der foreligger ikke oplysninger om alder af bygninger ellerstaldrenovering i de pågældende systemer.Resultater:Bredden af farestierne varierede mellem 115 og 248 cm (median 157 cm, 25-75 % fraktil:152-165 cm), mens længden af farestierne varierede mellem 200 og 315 cm (median 252cm, 25-75 % fraktil: 247-260 cm). Areal af farestien varierede mellem 2,85 og 6,15 m2(median 3,95 m2, 25-75 % fraktil: 3,75-4,25 m2).Andelen af spaltegulv i farestierne varierede mellem 22 og 100 % (median 53 %spaltegulv, 25-75 % fraktil: 51-58 % spaltegulv). I 11 af de 84 besætninger fandtes
76
PATTEGRISDØDELIGHED I DK
staldsektioner, hvor farestierne havde fuldspaltegulv. De almindeligst forekommendespalter i farestierne var i støbejern, som sås i 70/84 besætninger. Desuden havde 6/84besætninger sektioner med spaltegulve af beton, i 9/84 besætninger fandtes farestier medbåde spalter i beton samt støbejern eller metal, i 8/84 besætninger fandtes plastspalter ifarestier, mens 10/84 besætninger havde metalriste i farestierne.I 71/84 besætninger blev observeret sektioner med farestier uden strøelse. De hyppigstanvendte strøelsesmaterialer i farestalds-sektionerne var savsmuld og snittet halm. 7/84besætninger anvendte snittet halm, 11/84 besætninger anvendte savsmuld, 1 besætninganvendte både halm og savsmuld og 2/84 besætninger anvendte træmel.Søerne var fikseret i farebøjler hvis længde fra krybbe-bagkant varierede mellem 180 og220 cm (median 198 cm, 25-75 % fraktil 190-200 cm), mens bredden ved krybbensbagkant varierede mellem 48 og 143 cm (median 57 cm, 25-75 % fraktil: 55-60 cm). Hos3824 søer 2-4 uger efter faring i de 98 besætninger varierede bøjlebredden bagtil fra 42 til88 cm (median 64 cm, 25-75 % fraktil: 58-70 cm).Højden af skillevægge mellem farestier varierede fra 21 til 110 cm (median 53 cm, 25-75% fraktil: 52-55 cm).I 52/84 besætninger var der i en eller flere farestald-sektioner tydelig eller kraftig støj fraventilationssystemet, og i 2 af disse besætninger var der desuden tydelig støj fra radio. I9/84 besætninger var der tydelig eller kraftig støj fra andre støjkilder, som eksempelvisfoderanlæg, skrigende grise eller andre ikke angivne støjkilder.
77